Plasmodium
| Plasmodium | |
|---|---|
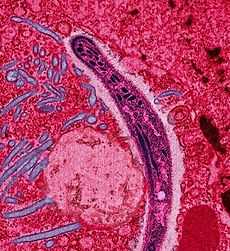 | |
| False-colored electron micrograph of a Plasmodium sp. sporozoite. | |
| Scientific classification | |
| Kingdom: | Chromalveolata |
| Superphylum: | Alveolata |
| Phylum: | Apicomplexa |
| Class: | Aconoidasida |
| Order: | Haemosporida |
| Family: | Plasmodiidae |
| Genus: | Plasmodium Marchiafava & Celli, 1885 |
| Species | |
Plasmodium, commonly known as the malaria parasite, is a large genus of parasitic protozoa. As with some other genera of clinically important microorganisms, the genus name also yields a common noun; thus species of the genus are known as plasmodia. Infection with plasmodia is known as malaria, a deadly disease widespread in the tropics.
The parasite always has two hosts in its life cycle: a mosquito vector and a vertebrate host. The life-cycle is very complex, involving a sequence of different stages both in the vector and the host. These stages include sporozoites which are injected by the mosquito vector into the host's blood; latent hypnozoites which may rest undetected in the liver for up to 30 years; merosomes and merozoites which infect the red cells (erythrocytes) of the blood; trophozoites which grow in the red cells, and schizonts which divide there, producing more merozoites which leave to infect more red cells; and male and female sexual forms, gametocytes, which are taken up by other mosquitoes. In the mosquito's midgut, the gametocytes develop into gametes which fertilize each other to form motile zygotes which escape the gut, only to grow into new sporozoites which move to the mosquito's salivary glands, from where they are injected into the mosquito's next host, infecting it and restarting the cycle.
The genus Plasmodium was first described in 1885. It now contains about 200 species divided into several subgenera; as of 2006 the taxonomy was shifting, and species from other genera are likely to be added to Plasmodium. At least ten species infect humans; other species infect other animals, including birds, reptiles and rodents, while 29 species infect non-human primates. The parasite is thought to have originated from Dinoflagellates, photosynthetic protozoa.
The most common forms of human malaria are caused by Plasmodium falciparum, P. vivax, P. knowlesi, and P. malariae. P. falciparum, common in sub-Saharan Africa, and P. knowlesi, common in Southeast Asia, are especially dangerous.
Taxonomy and host range
The genus Plasmodium was created in 1885 by Marchiafava and Celli and there are over 200 species recognized. New species continue to be described.[1]
As of 2006, the genus is in need of reorganization as it has been shown that parasites belonging to the genera Haemocystis and Hepatocystis appear to be closely related to Plasmodium. It is likely that other species such as Haemoproteus meleagridis will be included in this genus once it is revised.
Host range among the mammalian orders is non uniform. At least 29 species infect non-human primates; rodents outside the tropical parts of Africa are rarely affected; a few species are known to infect bats, porcupines and squirrels; carnivores, insectivores and marsupials are not known to act as hosts.
In 1898 Ronald Ross demonstrated the existence of Plasmodium in the wall of the midgut and salivary glands of a Culex mosquito. For this discovery he won the Nobel Prize in 1902. However credit must also be given to the Italian professor Giovanni Battista Grassi, who showed that human malaria could only be transmitted by Anopheles mosquitoes. For some species the vector may not be a mosquito.
Mosquitoes of the genera Culex, Anopheles, Culiseta, Mansonia and Aedes may act as vectors. The known vectors for human malaria (more than 100 species) belong to the genus Anopheles. Bird malaria is commonly carried by species belonging to the genus Culex. Only female mosquitoes bite. Aside from blood both sexes live on nectar, but one or more blood meals are needed by the female for egg laying, because there is very little protein in nectar.
Life cycle
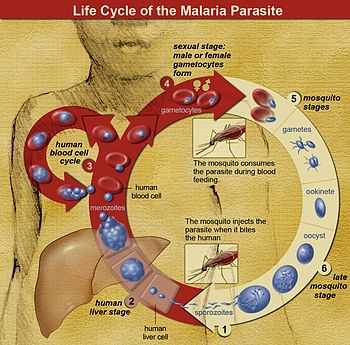
The life cycle of Plasmodium is very complex. Sporozoites from the saliva of a biting female mosquito are transmitted to either the blood or the lymphatic system[2] of the recipient. The sporozoites then migrate to the liver and invade hepatocytes. This latent or dormant stage of the Plasmodium sporozoite in the liver is called the hypnozoite.
The development from the hepatic stages to the erythrocytic stages has been obscure. In 2006[3] it was shown that the parasite buds off the hepatocytes in merosomes containing hundreds or thousands of merozoites. These merosomes have been subsequently shown[4] to lodge in the pulmonary capillaries and to disintegrate there slowly over 48–72 hours releasing merozoites. Erythrocyte invasion is enhanced when blood flow is slow and the cells are tightly packed: both of these conditions are found in the alveolar capillaries.
Within the erythrocytes the merozoites grow first to a ring-shaped form and then to a larger trophozoite form. In the schizont stage, the parasite divides several times to produce new merozoites, which leave the red blood cells and travel within the bloodstream to invade new red blood cells. Most merozoites continue this replicative cycle, but some merozoites differentiate into male or female sexual forms (gametocytes) (also in the blood), which are taken up by the female mosquito.
In the mosquito's midgut, the gametocytes develop into gametes and fertilize each other, forming a zygote. After a brief period of inactivity, zygotes transform into a motile form called ookinetes. The ookinetes penetrate and escape the midgut, then embed themselves onto the exterior of the gut membrane and transform into oocysts. The nuclei of oocysts divide many times to produce large numbers of tiny elongated sporozoites. These sporozoites migrate to the salivary glands of the mosquito where they are injected into the blood of the next host the mosquito bites. The sporozoites move to the liver where they repeat the cycle.
The pattern of alternation of sexual and asexual reproduction is common in parasitic species. The evolutionary advantages of this type of life cycle were recognised by Mendel. Under favourable conditions, asexual reproduction is superior to sexual as the parent is well adapted to its environment and its descendents share all its genes. Transferring to a new host or in times of stress, sexual reproduction is generally superior as it shuffles the genes of two parents, producing a variety of individuals, some of which will be better adapted to the new environment.
Reactivation of the hypnozoites has been reported for up to 30 years after the initial infection in humans. The factors precipating this reactivation are not known. In the species Plasmodium malariae, Plasmodium ovale and Plasmodium vivax hypnozoites have been shown to occur. Reactivation does not occur in infections with Plasmodium falciparum. It is not known if hypnozoite reactivaction may occur with any of the remaining species that infect humans but this is presumed to be the case.
Evolution
The life cycle of Plasmodium is best understood in terms of its evolution.
The Apicomplexa—the phylum to which Plasmodium belongs—are thought to have originated within the Dinoflagellates, a large group of photosynthetic protozoa. It is thought that the ancestors of the Apicomplexa were originally prey organisms that evolved the ability to invade the intestinal cells and subsequently lost their photosynthetic ability. Some extant Dinoflagellates, however, can invade the bodies of jellyfish and continue to photosynthesize, which is possible because jellyfish bodies are almost transparent. In other organisms with opaque bodies this ability would most likely rapidly be lost.
It is thought that Plasmodium evolved from a parasite spread by the fecal-oral route which infected the intestinal wall. At some point this parasite evolved the ability to infect the liver. This pattern is seen in the genus Cryptosporidium, to which Plasmodium is distantly related. At some later point this ancestor developed the ability to infect blood cells and to survive and infect mosquitoes. Plasmodium subsequently evolved a mechanism to invade the salivary glands of mosquitoes, allowing for transmission from mosquito to host. Once mosquito transmission was firmly established, the previous fecal-oral route was lost within the Plasmodium genus.[5]
The survivorship and relative fitness of mosquitoes are not adversely affected by Plasmodium infection which indicates the importance of vector fitness in shaping the evolution of Plasmodium.[6] Plasmodium has evolved the capability to manipulate mosquito feeding behavior. Mosquitoes harboring Plasmodium have a higher propensity to bite than uninfected mosquitoes. This tendency has facilitated the spread of Plasmodium to the various hosts. [7]
Current (2007) theory suggests that the genera Plasmodium, Hepatocystis and Haemoproteus evolved from Leukocytozoon species. Parasites of the genus Leukocytozoan infect white blood cells (leukocytes), liver and spleen cells and are transmitted by 'black flies' (Simulium species) — a large genus of flies related to the mosquitoes.
Leukocytes, hepatocytes and most spleen cells actively phagocytose particulate matter, making entry into the cell easier for the parasite. The mechanism of entry of Plasmodium species into erythrocytes is still very unclear, taking as it does less than 30 seconds. It is not yet known if this mechanism evolved before mosquitoes became the main vectors for transmission of Plasmodium.
Plasmodium evolved about 130 million years ago. This period coincided with the rapid spread of the angiosperms (flowering plants). This expansion in the angiosperms is thought to be due to at least one genomic duplication event. It seems probable that the increase in the number of flowers led to an increase in the number of mosquitoes and their contact with vertebrates.
Environmental factors play a considerable role in the evolution of Plasmodium and the transmission of malaria. The genetic information of Plasmodium falciparum has signaled a recent expansion that coincides with the agricultural revolution It is likely that the development of extensive agriculture increased mosquito population densities by giving rise to more breeding sites, which may have triggered the evolution and expansion of Plasmodium falciparum. [8]
Mosquitoes evolved in what is now South America about 230 million years ago. There are over 3500 species recognised but to date their evolution has not been well worked out so a number of gaps in our knowledge of the evolution of Plasmodium remain. It seems probable that birds were the first group infected by Plasmodium followed by the reptiles — probably the lizards. At some point primates and rodents became infected. The remaining species infected outside these groups seem likely to be due to relatively recent events.
There are over one hundred species of mosquito-transmitted Plasmodium. The phylogeny of these malarial parasites suggests that the Plasmodium of mammalian hosts forms a well-defined clade strongly associated with the specialization to the Anopheles mosquito vector. This was a major evolutionary transition that allowed Plasmodium to exploit humans and other mammals.[9]
P. falciparum, the most lethal malaria parasite of humans, evolved from a "nearly identical" parasite of western gorillas, not from chimpanzees, bonobos or ancient human populations.[10]
The high mortality and morbidity caused by malaria—especially that caused by P. falciparum —has placed the greatest selective pressure on the human genome in recent history. Several genetic factors provide some resistance to Plasmodium infection, including sickle cell trait, thalassaemia traits, glucose-6-phosphate dehydrogenase deficiency, and the absence of Duffy antigens on red blood cells.[11] [12]
Although there are therapeutic medications to treat malaria, Plasmodium has accumulated increasing drug resistance over time. A recent examination has shown that even artemisinin, one of the most powerful anti-malarial drugs, has been experiencing decreased efficacy due to the development of resistance. [13]
Molecular biology

All the species examined to date have 14 chromosomes, one mitochondrion and one plastid (also known as apicoplast, an organelle similar to a chloroplast). The chromosomes vary from 500 kilobases to 3.5 megabases in length. It is presumed that this is the pattern throughout the genus. The plastid, unlike those found in algae, is not photosynthetic. Its function is not fully known; however, it has been demonstrated that some essential metabolic pathways like isoprenoid, Fe-S clusters, fatty acid and phospholipid biosynthesis occurs in this organelle, and that it also still possess its own genome, partly shared with the nucleus.[14]
On a molecular level, the parasite damages red blood cells using plasmepsin enzymes — aspartic acid proteases which degrade hemoglobin.
Taxonomy
Plasmodium belongs to the family Plasmodiidae (Levine, 1988), order Haemosporidia and phylum Apicomplexa. There are 450 recognised species in this order. Many species of this order are undergoing reexamination of their taxonomy with DNA analysis. It seems likely that many of these species will be reassigned after these studies have been completed.[15][16] For this reason the entire order is outlined here.
- Genus Plasmodium
- Subgenus Asiamoeba (lizards)
- Subgenus Bennettinia (birds)
- Subgenus Carinamoeba (reptiles)
- Subgenus Giovannolaia (birds)
- Subgenus Haemamoeba (birds)
- Subgenus Huffia (birds)
- Subgenus Lacertamoeba (reptiles)
- Subgenus Laverania (higher primates)
- Subgenus Novyella (birds)
- Subgenus Paraplasmodium (lizards)
- Subgenus Plasmodium (monkeys, higher primates)
- Subgenus Sauramoeba (reptiles)
- Subgenus Vinckeia (non-primate mammals)
- Genus Polychromophilus
- Genus Rayella
- Genus Saurocytozoon
The genera Plasmodium, Fallisia and Saurocytozoon all cause malaria in lizards. All are carried by Diptera (true two-winged flies). Pigment is absent in the Garnia. Non pigmented gametocytes are typically the only forms found in Saurocytozoon: pigmented forms may be found in the leukocytes occasionally. Fallisia produce non pigmented asexual and gametocyte forms in leukocytes and thrombocytes.
Subgenera
The full taxonomic name of a species includes the subgenus but this is often omitted. The full name indicates some features of the morphology and type of host species.
The only two species in the sub genus Laverania are P. falciparum and P. reichenowi.
Species infecting monkeys and apes (the higher primates) with the exceptions of P. falciparum and P. reichenowi are classified in the subgenus Plasmodium.
Parasites infecting other mammals including lower primates (lemurs and others) are classified in the subgenus Vinckeia. The distinction between P. falciparum and P. reichenowi and the other species infecting higher primates was based on morphological findings but have since been confirmed by DNA analysis. Vinckeia, while previously considered to be something of a taxonomic 'rag bag', has been recently shown to form a coherent grouping. The remaining groupings here are based on the morphology of the parasites. Revisions to this system are likely as more species are subject to DNA analysis.
The four subgenera Giovannolaia, Haemamoeba, Huffia and Novyella were created by Corradetti et al.[17] for the known avian malarial species. A fifth — Bennettinia — was created in 1997 by Valkiunas.[18] The relationships between the subgenera are a matter of current investigation. Martinsen et al. 's recent (2006) paper outlines what was known at the time.[19]
As of 2007, P. juxtanucleare is the only known member of the subgenus Bennettinia.
Unlike the mammalian and bird malarias those affecting reptiles have been more difficult to classify. In 1966 Garnham classified those with large schizonts as Sauramoeba, those with small schizonts as Carinamoeba and the single then known species infecting snakes (Plasmodium wenyoni) as Ophidiella.[20] He was aware of the arbitrariness of this system and that it might not prove to be biologically valid. Telford in 1988 used this scheme as the basis for the accepted (2007) system.[21]
| Classification criteria |
|---|
| Avian species:
Species in the subgenus Bennettinia have the following characteristics:
Species in the subgenus Giovannolaia have the following characteristics:
Species in the subgenus Haemamoeba have the following characteristics:
Species in the subgenus Huffia have the following characteristics:
Species in the subgenus Novyella have the following characteristics:
Reptile species: Species in the subgenus Carinamoeba infect lizards. Their schizonts normally give rise to less than 8 merozoites, unlike those in the subgenus Sauramoeba which also infect lizards, but whose schizonts normally give rise to more than 8 merozoites.
Ophidiella was a subgenus created by Garnham in 1966 for the species infecting snakes. As of 2007 it was no longer in use. |
Species infecting humans

The species of Plasmodium that infect humans include:
- Plasmodium falciparum (the cause of malignant tertian malaria)
- Plasmodium vivax (the most frequent cause of benign tertian malaria)
- Plasmodium ovale (the other, less frequent, cause of benign tertian malaria)
- Plasmodium malariae (the cause of benign quartan malaria)
- Plasmodium knowlesi (the cause of severe quotidian malaria in South East Asia since 1965)
- Plasmodium brasilianum
- Plasmodium cynomolgi
- Plasmodium cynomolgi bastianellii
- Plasmodium inui
- Plasmodium rhodiani
- Plasmodium schweitzi
- Plasmodium semiovale
- Plasmodium simium
The first four listed here are the most common species that infect humans. Nearly all human deaths from malaria are caused by the first species, P. falciparum, mainly in sub-Saharan Africa. With the use of the polymerase chain reaction additional species have been and are still being identified that infect humans.
One possible experimental infection has been reported with Plasmodium eylesi. Fever and low grade parasitemia were apparent at 15 days. The volunteer (Dr Bennett) had previously been infected by Plasmodium cynomolgi and the infection was not transferable to a gibbon (P. eylesi 's natural host) so this cannot be regarded as definitive evidence of its ability to infect humans. A second case has been reported that may have been a case of P. eylesi but the author was not certain of the infecting species.[22]
A possible infection with Plasmodium tenue has been reported.[23] This report described a case of malaria in a three-year-old black girl from Georgia, US, who had never been outside the US. She suffered from both P. falciparum and P. vivax malaria and while forms similar to those described for P. tenue were found in her blood even the author was skeptical about the validity of the diagnosis.
Confusingly, P. tenue was proposed in the same year (1914) for a species found in birds. The human species is now considered probably a misdiagnosis, and the bird species is described on the P. tenue page.
The only known host of P. falciparum and P. malariae is humans. P. vivax however can infect chimpanzees. Infection tends to be low grade but may be persistent and remain as source of parasites for humans for some time. P. vivax can also infect orangutans.[24]
P. ovale can be transmitted to chimpanzees. P. ovale has an unusual distribution, being found in Africa, the Philippines and New Guinea. In spite of its admittedly poor transmission to chimpanzees given its discontigous spread, it is suspected that P. ovale is a zoonosis with an as yet unidentified host. If so, the host is likely to be a primate. The remaining species capable of infecting humans all have other primate hosts.
Plasmodium shortii and Plasmodium osmaniae are now considered junior synonyms of Plasmodium inui
Taxonomy in parasitology before DNA based methods was always problematic, and revisions are continuing, leaving many obsolete names for Plasmodium species that infect humans.[25]
| Obsolete names for Plasmodium species infecting humans |
|---|
| P. camerense P. causiasium |
Infections in primates

The species that infect primates other than humans include: P. bouillize, P. brasilianum, P. bucki, P. cercopitheci,P. coatneyi, P. coulangesi, P. cynomolgi, P. eylesi, P. fieldi, P. foleyi, P. fragile, P. girardi, P. georgesi, P. gonderi, P. hylobati, P. inui, P. jefferyi, P. joyeuxi, P. knowlesi, P. lemuris, P. percygarnhami, P. petersi, P. reichenowi, P. rodhaini, P. sandoshami, P. semnopitheci, P. silvaticum, P. simiovale, P. simium, P. uilenbergi, P. vivax and P. youngei.
Most if not all Plasmodium species infect more than one host: the host records shown here should be regarded as incomplete.
| Host records |
|---|
*P. bouillize — Cercopithecis campbelli
|
| Primate mosquito vectors |
|---|
*Anopheles albimanus — P. vivax
|
| Subspecies infecting primates |
|---|
*P. cynomolgi — P. cynomolgi bastianelli and P. cynomolgi ceylonensis.
The evolution of these species is still being worked out and the relationships given here should be regarded as tentative. This grouping, while originally made on morphological grounds, now has considerable support at the DNA level.
|
Infections in non-primate mammals

The subgenus Vinckeia was created by Garnham to accommodate the mammalian parasites other than those infecting primates. Species infecting lemurs have also been included in this subgenus.
P. aegyptensis, P. bergei, P. chabaudi, P. inopinatum, P. yoelli and P. vinckei infect rodents. P. bergei, P. chabaudi, P. yoelli and P. vinckei have been used to study malarial infections in the laboratory. Other members of this subgenus infect other mammalian hosts.
| Host records |
|---|
*P. aegyptensis — Egyptian grass rat (Arvicanthis noloticus)[31]
|
| Vectors |
|---|
|
| Subspecies |
|---|
*P. berghei — P. berghei yoelii
Calomys callosus seems unlikely to be a natural host for P. atheruri, as P. atheruri is found in Africa and Calomys callosus in South America. |
| Less well documented species |
|---|
|
The species listed here from Courtney et al.[25] should be regarded as dubious.
P. epomophori of the bat (Hypsignathus monstruosus) has been reclassified as Hepatocystis epomophori. |
Infections in birds

Species in five Plasmodium subgenera infect birds — Bennettinia, Giovannolaia, Haemamoeba, Huffia and Novyella.[36] Giovannolaia appears to be a polyphyletic group and may be sudivided in the future.[19] DNA evidence is in 2014 helping to improve understanding of the diversity of Plasmodium species that infect birds.[37]
| Avian parasites |
|---|
| Species infecting birds include: P. accipiteris, P. alloelongatum, P. anasum, P. ashfordi, P. bambusicolai, P. bigueti, P. biziurae, P. buteonis, P. cathemerium, P. circumflexum, P. coggeshalli, P. corradettii, P. coturnix, P. dissanaikei, P. durae, P. elongatum, P. fallax, P forresteri, P. gallinacium, P. garnhami, P. giovannolai, P. griffithsi, P. gundersi, P. guangdong, P. hegneri, P. hermani, P. hexamerium, P. huffi, P. jiangi, P. juxtanucleare, P. kempi, P. lophurae, P.lutzi, P. matutinum, P. nucleophilum, P. papernai, P. paranucleophilum, P. parvulum, P. pediocetti, P. paddae, P. pinotti, P. polare, P. relictum, P. rouxi, P. tenue, P. tejerai, P. tumbayaensis and P. vaughani. |
| Avian host records |
|---|
*P. accipiteris — Levant sparrowhawk (Accipiter brevipes)
|
| Avian malaria vectors |
|---|
|
Aedes species:
Culex species:
Mansonia species:
|
| Avian malaria subspecies |
|---|
|
| Avian malaria inter-relatedness and doubtful species |
|---|
*P. durae is related to P. asanum, P. circumflexum, P. fallax, P. formosanum, P. gabaldoni, P. hegneri, P. lophrae, P. lophrae, P. pediocetti, P. pinotti, and P. polare.
A number of additional species have been described in birds — P. centropi, P. chloropsidis, P. gallinuae, P. herodialis, P. heroni, P. mornony, P. pericorcoti and P. ploceii — but the suggested speciation was based at least in part on the idea — 'one host — one species'. It has not been possible to reconcile the descriptions with any of the recognised species and these are not regarded as valid species. As further investigations are made into this genus these species may be resurrected. A species P. japonicum has been reported[48] but this appears to be the only report of this species and it should therefore be regarded of dubious validity. |
Infections in reptiles

Species in the subgenera Asiamoeba, Carinamoeba, Lacertaemoba, Paraplasmodium and Sauramoeba infect reptiles.[49]
Over 90 species and subspecies of Plasmodium infect lizards and they have been reported from over 3200 species of lizard and 29 species of snake. Only three species — P. pessoai, P. tomodoni and P. wenyoni — infect snakes.
| Species infecting reptiles |
|---|
| P. achiotense, P. aeuminatum, P. agamae, P. arachniformis, P. attenuatum,P. aurulentum, P. australis, P. azurophilum, P. balli, P. basilisci, P. beebei, P. beltrani , P. brumpti, P. brygooi, P. chiricahuae, P. circularis, P. cnemaspi, P. cnemidophori, P. colombiense, P. cordyli, P. diminutivum, P. diploglossi, P. egerniae, P. fairchildi, P. floridense, P. gabaldoni, P. giganteum, P. gologoense, P. gracilis, P. guyannense, P. heischi, P. holaspi, P. icipeensis, P. iguanae, P. josephinae, P. kentropyxi, P. lacertiliae, P. lainsoni, P. lepidoptiformis, P. lionatum, P. loveridgei, P. lygosomae, P. mabuiae, P. mackerrasae, P. maculilabre, P. marginatum, P. mexicanum, P. michikoa, P. minasense, P. pelaezi, P. pessoai, P. pifanoi, P. pitmani, P. rhadinurum, P. sasai,P. saurocaudatum, P. scorzai, P. siamense, P. robinsoni, P. sasai, P. scorzai, P. tanzaniae, P. tomodoni, P. torrealbai, P. tribolonoti, P. tropiduri, P. uluguruense, P. uzungwiense, P. vacuolatum, P. vastator, P. volans, P. wenyoni and P. zonuriae. |
| Host records |
|---|
*P. agamae — the rainbow lizard (Agama agama)
|
| Vectors |
|---|
*P. agamae — Lutzomyia or Culicoides species
|
| Subspecies |
|---|
*P. fairchildi — P. fairchildi fairchildi and P. fairchildi hispaniolae
|
| Inter-relatedness |
|---|
| *P. floridense is closely related to P. tropiduri and P. minasense |
Species reclassified into other genera
| Reclassified species |
|---|
As of 2007 the following species are regarded as belonging to the genus Hepatocystis rather than Plasmodium.
|
References
- ↑ Chavatte JM, Chiron F, Chabaud A, Landau I (March 2007). "[Probable speciations by "host-vector 'fidelity'": 14 species of Plasmodium from magpies]". Parasite (in French) 14 (1): 21–37. PMID 17432055.
- ↑ "Malaria Parasites Develop in Lymph Nodes". HHMI News. Howard Hughes Medical Institute. 22 January 2006.
- ↑ Sturm A, Amino R, van de Sand C et al. (September 2006). "Manipulation of host hepatocytes by the malaria parasite for delivery into liver sinusoids". Science 313 (5791): 1287–90. doi:10.1126/science.1129720. PMID 16888102.
- ↑ Baer K, Klotz C, Kappe SH, Schnieder T, Frevert U (November 2007). "Release of hepatic Plasmodium yoelii merozoites into the pulmonary microvasculature". PLoS Pathog. 3 (11): e171. doi:10.1371/journal.ppat.0030171. PMC 2065874. PMID 17997605.
- ↑ Ghosh, A.K; Devenport, M; Jethwaney, D. "Malaria parasite invasion of the mosquito salivary glands requires interaction between the Plasmoidum TRAP and the Anopheles saglin proteins". PLoS Pathog. 5(1):e1000265.
- ↑ Ferguson, HM; Mackinnon, MJ; Chan, BH; Read, AF. (2003). "Mosquito mortality and the evolution of malaria virulence". Evolution 57 (12): 2792–804. doi:10.1554/03-211.
- ↑ Koella, J.C.; Sørensen, F.L.; Anderson, R.A. (1998). "The malaria parasite, Plasmodium falciparum, increases the frequency of multiple feeding of its mosquito vector, Anopheles gambiae". Proc Biol Sci 265 (1398): 763–8. doi:10.1098/rspb.1998.0358.
- ↑ Hume J.C., Lyons E.J., Day K.P. 2003. Human migration, mosquitoes and the evolution of Plasmodium falciparum. Trends Parasitol. 19(3):144-9.
- ↑ Martinsen ES, Perkins SL, Schall JJ. (2008). "A three-genome phylogeny of malaria parasites (Plasmodium and closely related genera): evolution of life-history traits and host switches". Molecular Phylogenetic Evolution 47 (1): 261–273. doi:10.1016/j.ympev.2007.11.012. PMID 18248741.
- ↑ Weimin Liu, Yingying Li, Gerald H. Learn, Rebecca S. Rudicell, Joel D. Robertson, Brandon F. Keele, Jean-Bosco N. Ndjango, Crickette M. Sanz, David B. Morgan, Sabrina Locatelli, Mary K. Gonder, Philip J. Kranzusch, Peter D. Walsh, Eric Delaporte, Eitel Mpoudi-Ngole, Alexander V. Georgiev, Martin N. Muller, George M. Shaw, Martine Peeters, Paul M. Sharp, Julian C. Rayner & Beatrice H. Hahn (23 September 2010). "Origin of the human malaria parasite Plasmodium falciparum in gorillas" 467 (7314). pp. 420–425. doi:10.1038/nature09442. PMC 2997044. PMID 20864995.
- ↑ Kwiatkowski DP (2005). "How malaria has affected the human genome and what human genetics can teach us about malaria". American Journal of Human Genetics 77 (2): 171–92. doi:10.1086/432519. PMC 1224522. PMID 16001361.

- ↑ Hedrick PW (2011). "Population genetics of malaria resistance in humans". Heredity 107 (4): 283–304. doi:10.1038/hdy.2011.16. PMC 3182497. PMID 21427751.
- ↑ Ashley, E.A.; Dhorda, M.; Fairhurst, R.M. et al. (2014). "Spread of artemisinin resistance in Plasmodium falciparum malaria". N Engl J Med 371 (5): 411–23. doi:10.1056/nejmoa1314981.
- ↑ vanDooren, G; Striepen, B (2013). "The Algal Past and Parasite Present of Apicoplast". Annual Reviews of Microbiology 67: 271–289. doi:10.1146/annurev-micro-092412-155741. PMID 23808340.
- ↑ Perkins SL, Schall JJ (October 2002). [0972:AMPOMP2.0.CO;2 "A molecular phylogeny of malarial parasites recovered from cytochrome b gene sequences"]. J. Parasitol. 88 (5): 972–8. doi:10.1645/0022-3395(2002)088[0972:AMPOMP]2.0.CO;2. PMID 12435139.
- ↑ Yotoko, K.S.C.; Elisei C. (November 2006). "Malaria parasites (Apicomplexa, Haematozoea) and their relationships with their hosts: is there an evolutionary cost for the specialization?". J. Zoo. Syst. Evol. Res. 44 (4): 265–273. doi:10.1111/j.1439-0469.2006.00377.x.
- ↑ Corradetti A., Garnham P.C.C., Laird M. (1963). "New classification of the avian malaria parasites". Parassitologia 5: 1–4.
- ↑ Valkiunas G (1997). "Bird Haemosporidia". Acta Zoologica Lituanica 3–5: 1–607. ISSN 1392-1657.
- ↑ 19.0 19.1 19.2 Martinsen ES, Waite JL, Schall JJ (April 2007). "Morphologically defined subgenera of Plasmodium from avian hosts: test of monophyly by phylogenetic analysis of two mitochondrial genes". Parasitology 134 (Pt 4): 483–90. doi:10.1017/S0031182006001922. PMID 17147839.
- ↑ Garnham 1966
- ↑ Telford S (1988). "A contribution to the systematics of the reptilian malaria parasites, family Plasmodiidae (Apicomplexa: Haemosporina)". Bulletin of the Florida State Museum Biological Sciences 34 (2): 65–96.
- ↑ Tsukamoto M (1977). "An imported human malarial case characterized by severe multiple infections of the red blood cells". Ann. Trop. Med. Parasit. 19 (2): 95–104.
- ↑ Russel P.F. (1928). "Plasmodium tenue (Stephens): A review of the literature and a case report". Am. J. Trop. Med. s1–8 (5): 449–479.
- ↑ Reid MJ, Ursic R, Cooper D et al. (December 2006). "Transmission of human and macaque Plasmodium spp. to ex-captive orangutans in Kalimantan, Indonesia". Emerging Infect. Dis. 12 (12): 1902–8. doi:10.3201/eid1212.060191. PMC 3291341. PMID 17326942.
- ↑ 25.0 25.1 Coatney G.R., Roudabush R.L. (1936). "A catalog and host-index of the genus Plasmodium". J. Parasitol. 22 (4): 338–353. doi:10.2307/3271859.
- ↑ Collins WE, Sullivan JS, Nace D, Williams T, Williams A, Barnwell JW (February 2008). "Observations on the sporozoite transmission of Plasmodium vivax to monkeys". J. Parasitol. 94 (1): 287–8. doi:10.1645/GE-1283.1. PMID 18372652.
- ↑ Collins WE, Richardson BB, Morris CL, Sullivan JS, Galland GG (July 1998). "Salvador II strain of Plasmodium vivax in Aotus monkeys and mosquitoes for transmission-blocking vaccine trials". Am. J. Trop. Med. Hyg. 59 (1): 29–34. PMID 9684622.
- ↑ Collins WE, Sullivan JS, Nace D et al. (April 2002). [0295:EIOAFW2.0.CO;2 "Experimental infection of Anopheles farauti with different species of Plasmodium"]. J. Parasitol. 88 (2): 295–8. doi:10.1645/0022-3395(2002)088[0295:EIOAFW]2.0.CO;2. PMID 12054000.
- ↑ Collins WE, Morris CL, Richardson BB, Sullivan JS, Galland GG (August 1994). "Further studies on the sporozoite transmission of the Salvador I strain of Plasmodium vivax". J. Parasitol. 80 (4): 512–7. doi:10.2307/3283184. PMID 8064516.
- ↑ Tan CH, Vythilingam I, Matusop A, Chan ST, Singh B (2008). "Bionomics of Anopheles latens in Kapit, Sarawak, Malaysian Borneo in relation to the transmission of zoonotic simian malaria parasite Plasmodium knowlesi". Malar. J. 7: 52. doi:10.1186/1475-2875-7-52. PMC 2292735. PMID 18377652.
- ↑ Abd-el-Aziz GA, Landau I, Miltgen F (1975). "[Description of Plasmodium aegyptensis n. sp., presumed parasite of the Muridae Arvicanthis noloticus in Upper Egypt]". Ann Parasitol Hum Comp (in French) 50 (4): 419–24. PMID 1211772.
- ↑ Sandosham AA, Yap LF, Omar I (September 1965). "A malaria parasite, plasmodium (Vinckeia) booliati sp.nov., from a Malayan giant flying squirrel". Med J Malaya 20 (1): 3–7. PMID 4221411.
- ↑ Keymer IF (June 1966). "Studies on Plasmodium (Vinckeia) cephalophi of the grey duiker (Sylvicapra grimmia)". Ann Trop Med Parasitol 60 (2): 129–38. PMID 5962467.
- ↑ Landau I, Chabaud AG (1978). "[Description of P. cyclopsi n. sp. a parasite of the microchiropteran bat Hipposideros cyclops in Gabon (author's transl)]". Ann Parasitol Hum Comp (in French) 53 (3): 247–53. PMID 697287.
- ↑ Lien JC, Cross JH (December 1968). "Plasmodium (Vinckeia) watteni sp. n. from the Formosan giant flying squirrel, Petaurista petaurista grandis". J. Parasitol. 54 (6): 1171–4. doi:10.2307/3276986. PMID 5757690.
- ↑ Wiersch SC, Maier WA, Kampen H (May 2005). "Plasmodium (Haemamoeba) cathemerium gene sequences for phylogenetic analysis of malaria parasites". Parasitol. Res. 96 (2): 90–4. doi:10.1007/s00436-005-1324-8. PMID 15812672.
- ↑ Clark, Nicholas; Clegg, Sonya; Lima, Marcos (2014). "A review of global diversity in avian haemosporidians (Plasmodium and Haemoproteus: Haemosporida): new insights from molecular data". International Journal for Parasitology 44 (5): 329–338. doi:10.1016/j.ijpara.2014.01.004. PMID 24556563.
- ↑ Valkiūnas G, Zehtindjiev P, Hellgren O, Ilieva M, Iezhova TA, Bensch S (May 2007). "Linkage between mitochondrial cytochrome b lineages and morphospecies of two avian malaria parasites, with a description of Plasmodium (Novyella) ashfordi sp. nov". Parasitol. Res. 100 (6): 1311–22. doi:10.1007/s00436-006-0409-3. PMID 17235548.
- ↑ Landau I, Chabaud AG, Bertani S, Snounou G (December 2003). "Taxonomic status and re-description of Plasmodium relictum (Grassi et Feletti, 1891), Plasmodium maior Raffaele, 1931, and description of P. bigueti n. sp. in sparrows". Parassitologia 45 (3-4): 119–23. PMID 15267099.
- ↑ Kirkpatrick CE, Lauer DM (January 1985). "Hematozoa of raptors from southern New Jersey and adjacent areas". J. Wildl. Dis. 21 (1): 1–6. doi:10.7589/0090-3558-21.1.1. PMID 3981737.
- ↑ Earlé RA, Horak IG, Huchzermeyer FW, Bennett GF, Braack LE, Penzhorn BL (September 1991). "The prevalence of blood parasites in helmeted guineafowls, Numida meleagris, in the Kruger National Park". Onderstepoort J. Vet. Res. 58 (3): 145–7. PMID 1923376.
- ↑ Valkiūnas G, Zehtindjiev P, Dimitrov D, Krizanauskiene A, Iezhova TA, Bensch S (May 2008). "Polymerase chain reaction-based identification of Plasmodium (Huffia) elongatum, with remarks on species identity of haemosporidian lineages deposited in GenBank". Parasitol. Res. 102 (6): 1185–93. doi:10.1007/s00436-008-0892-9. PMID 18270739.
- ↑ Murata K, Nii R, Sasaki E et al. (February 2008). "Plasmodium (Bennettinia) juxtanucleare infection in a captive white eared-pheasant (Crossoptilon crossoptilon) at a Japanese zoo". J. Vet. Med. Sci. 70 (2): 203–5. doi:10.1292/jvms.70.203. PMID 18319584.
- ↑ Christensen BM, Barnes HJ, Rowley WA (July 1983). "Vertebrate host specificity and experimental vectors of Plasmodium (Novyella) kempi sp. n. from the eastern wild turkey in Iowa". J. Wildl. Dis. 19 (3): 204–13. doi:10.7589/0090-3558-19.3.204. PMID 6644918.
- ↑ Manwell RD (November 1968). "Plasmodium octamerium n. sp., an avian malaria parasite from the pintail whydah bird Vidua macroura". J. Protozool. 15 (4): 680–5. doi:10.1111/j.1550-7408.1968.tb02194.x. PMID 5719065.
- ↑ Valkiũnas G, Iezhova TA (August 2001). [0930:ACOTBP2.0.CO;2 "A comparison of the blood parasites in three subspecies of the yellow wagtail Motacilla flava"]. J. Parasitol. 87 (4): 930–4. doi:10.1645/0022-3395(2001)087[0930:ACOTBP]2.0.CO;2. PMID 11534666.
- ↑ Poinar G (May 2005). "Plasmodium dominicana n. sp. (Plasmodiidae: Haemospororida) from Tertiary Dominican amber". Syst. Parasitol. 61 (1): 47–52. doi:10.1007/s11230-004-6354-6. PMID 15928991.
- ↑ Manwell RD (February 1966). "Plasmodium japonicum, P. juxtanucleare and P. nucleophilum in the Far East". J. Protozool. 13 (1): 8–11. doi:10.1111/j.1550-7408.1966.tb01860.x. PMID 5912391.
- ↑ Schall JJ (December 2000). "Transmission success of the malaria parasite Plasmodium mexicanum into its vector: role of gametocyte density and sex ratio". Parasitology 121 (Pt 6): 575–80. doi:10.1017/s0031182000006818. PMID 11155927.
- ↑ Southgate BA (1970). "Plasmodium (Sauramoeba) giganteum in Agama cyanogaster: a new host record". Trans. R. Soc. Trop. Med. Hyg. 64 (1): 12–3. PMID 5462484.
- ↑ Garnham PC, Telford SR (November 1984). "A new malaria parasite Plasmodium (Sauramoeba) heischi in skinks (Mabuya striata) from Nairobi, with a brief discussion of the distribution of malaria parasites in the family Scincidae". J. Protozool. 31 (4): 518–21. doi:10.1111/j.1550-7408.1984.tb05494.x. PMID 6512723.
- ↑ Telford SR (October 1986). "Fallisia parasites (Haemosporidia: Plasmodiidae) from the flying lizard, Draco maculatus (Agamidae) in Thailand". J. Parasitol. 72 (5): 766–9. doi:10.2307/3281471. PMID 3100759.
- ↑ 53.0 53.1 Telford SR (1979). "A taxonomic revision of small neotropical saurian Malarias allied to Plasmodium minasense". Ann Parasitol Hum Comp 54 (4): 409–22. PMID 533109.
- ↑ Telford SR, Telford SR (April 2003). [0362:RAROPP2.0.CO;2 "Rediscovery and redescription of Plasmodium pifanoi and description of two additional Plasmodium parasites of Venezuelan lizards"]. J. Parasitol. 89 (2): 362–8. doi:10.1645/0022-3395(2003)089[0362:RAROPP]2.0.CO;2. PMID 12760655.
Further reading
Identification
- Garnham, P.C.C. (1966). Malaria Parasites And Other Haemosporidia. Oxford: Blackwell. ISBN 0397601328.
- Hewitt, R.I. (1940). Bird Malaria. American Journal of Hygiene 15. Baltimore: Johns Hopkins Press.
- Laird, M. (1998). Avian Malaria in the Asian Tropical Subregion. Singapore: Springer. ISBN 9813083190.
Biology
- Baldacci P, Ménard R (October 2004). "The elusive malaria sporozoite in the mammalian host". Mol. Microbiol. 54 (2): 298–306. doi:10.1111/j.1365-2958.2004.04275.x. PMID 15469504.
- Bledsoe GH (December 2005). "Malaria primer for clinicians in the United States" (PDF). South. Med. J. 98 (12): 1197–204; quiz 1205, 1230. doi:10.1097/01.smj.0000189904.50838.eb. PMID 16440920.
- Shortt HE (1951). "Life-cycle of the mammalian malaria parasite". Br. Med. Bull. 8 (1): 7–9. PMID 14944807.
History
- Slater LB (2005). "Malarial birds: modeling infectious human disease in animals". Bull Hist Med 79 (2): 261–94. doi:10.1353/bhm.2005.0092. PMID 15965289.
External links
| Wikispecies has information related to: Plasmodium |
- Malaria Atlas Project
- "Plasmodium". NCBI Taxonomy Browser. 5820.