Peptidomimetic
A peptidomimetic is a small protein-like chain designed to mimic a peptide. They typically arise either from modification of an existing peptide, or by designing similar systems that mimic peptides, such as peptoids and β-peptides. Irrespective of the approach, the altered chemical structure is designed to advantageously adjust the molecular properties such as, stability or biological activity. This can have a role in the development of drug-like compounds from existing peptides. These modifications involve changes to the peptide that will not occur naturally (such as altered backbones and the incorporation of nonnatural amino acids).
D-peptide peptidomimetics
A D-peptide is a small sequence of D-amino acids. Since ribosomes are specific to L-amino acids, D-peptides rarely occur naturally in organisms and are not easily digested or degraded. D-peptide peptidomimetics are D-peptides designed to mimic natural L-peptides that commonly have therapeutic properties.
Properties of D-peptides
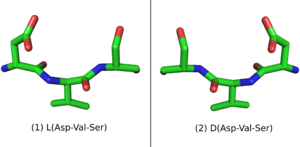
When placed in a nonchiral solvent like water, D-peptides, as well as the larger polypeptide D-proteins, have similar but mirrored properties to the L-peptides and L-proteins with identical sequences. If an L-protein does not require a Chaperone or a structural cofactor to fold, its D-enantiomer protein should have a mirror image conformation with respect to the L-protein (Figure 1). A D-enzyme should act on substrates of reverse chirality compared to the L-enzyme with the same sequence. Similarly, if an L-peptide binds to an L-protein, their D-peptide and D-protein counterparts should bind together in a mirrored way.[1]
D-peptides also have properties that make them attractive as drugs. D-peptides are less susceptible to be degraded in the stomach or inside cells by proteolysis. D-peptide drugs can therefore be taken orally and are effective for a longer period of time. D-peptides are easy to synthesize, when compared to many other drugs. In some cases, D-peptides can have a low immunogenic response.[2]
Methods to design D-peptides
Ret design
An L-peptide has three analogue sequences (Figure 2) built from L and D amino acids: the D-enantiomer or inverso-peptide with the same sequence, but composed of D-amino acids and a mirror conformation; the retro-peptide, consisting of the same sequence of L amino acids but in reverse order; and the retro-inverso or D-retro-enantiomer peptide, consisting of D-amino acids in the reversed sequence.[3] [4]
While the L-peptide and its D-enantiomer are mirror structures of each other, the L-retro-peptide is the mirror image of the D-retro-inverso-peptide. On the other hand, the L-peptide and the D-retro-inverso-peptide share a similar arrangement of side-chains, although their carboxyl and amino groups point in opposing directions. For small peptides that do not depend on a secondary structure for binding, an L-peptide and its D-retro-inverso-peptide is likely to have a similar binding affinity with a target L-protein.

Mirror-image phage display
Phage display is a technique to screen large libraries of peptides for binding to a target protein. In phage display, the DNA sequence that codes the potential drug-peptide is fusioned to the gene of the protein coat of bacteriophages and introduced into a vector. Diversity can be introduced to the peptide by mutagenesis. The protein coats peptides are then expressed and purified, and applied to a surface of immobilized protein targets. The surface is then washed away to remove non-binding peptides, while the remaining binding peptides are eluted. [5]
Mirror-image phage display is a similar method that can be used to screen large libraries of D-peptides that bind to target L-proteins. More precisely, since D-peptides can not be expressed in bacteriophages, mirror-image phage display screens L-peptides that bind to immobilized D-proteins that are previously chemically synthesized. Because of the mirror properties of D-peptides, the D-enantiomer of an L-peptide that binds to a D-protein will bind to the L-protein.
Mirror-image phage display, however, has two disadvantages when compared to phage display. Target D-proteins must be chemically synthesized, which is normally an expensive and time consuming process. Also, the target protein must not require a cofactor or a chaperone to fold, otherwise the chemically synthesized D-protein will not fold to the target, mirror structure.

Examples
An example of peptidomimetics were those designed and synthesized with the purpose of binding to target proteins in order to induce cancer cells into a form of programmed cell death called apoptosis. Basically, these work by mimicking key interactions that activate apoptotic pathway in the cell.
All healthy cells in metazoa (multi-celled animals) are subject to programmed cell death when they are no longer wanted; but cancer cells have the ability to evade apoptosis and the body's attempts to get rid of them. So peptidomimetics are part of the wide effort by researchers, research labs and institutions to create cures for cancer by means of restoring or activating apoptotic pathways in specific cells.
An impressive advance in this effort was reported by Loren D. Walensky and colleagues in the 3 September 2004 issue of the journal Science.[6] This team succeeded in reducing a variety of human leukemic cells that had been xenografted (transplanted) into mice. The peptidomimetic that they succeeded in synthesizing, SAHB (stabilized alpha-helix of BCL-2 domains) specifically activates the mitochondrial apoptotic pathway in the aforementioned malignant cells (see intrinsic and extrinsic inducers of the apoptotic process), without harming healthy tissues.
Harran and co-workers reported another impressive advance in the same issue of Science.[7] They were able to synthesize a molecule that mimics the proapoptotic protein called SMAC (whose function is described in biochemical execution of apoptosis).
See also
- Apoptosis
- Cancer
- Beta-peptide
- Foldamers
- Non-proteinogenic amino acids
- Expanded genetic code
- Clicked peptide polymer
References
- ↑ Milton RC, Milton SC, Kent SB (1992). "Total chemical synthesis of a D-enzyme: the enantiomers of HIV-1 protease show demonstration of reciprocal chiral substrate specificity". Science 256: 1445–1448. doi:10.1126/science.1604320.
- ↑ Welch BD, VanDemark AP, Heroux A, Hill CP, Kay MS. (2007). "Potent D-peptide inhibitors of HIV-1 entry.". Proceedings of the National Academy of Sciences 104 (43): 16828–16833. doi:10.1073/pnas.0708109104. PMC 2040420. PMID 17942675.
- ↑ Guichard G, Benkirane N, Zeder-Lutz G, van Regenmortel MH, Briand JP, Muller S. (1994). "Antigenic mimicry of natural L-peptides with retro-inverso-peptidomimetics". Proceedings of the National Academy of Sciences 91 (21): 9765–9769. doi:10.1073/pnas.91.21.9765.
- ↑ Cardo-Vila M, Giordano RJ, Sidman RL, Bronk LF, Fan Z, Mendelsohn J, Arap W, Pasqualini R. (2010). "From combinatorial peptide selection to drug prototype (II): Targeting the epidermal growth factor receptor pathway.". Proceedings of the National Academy of Sciences 107 (11): 5118–5123. doi:10.1073/pnas.0915146107. PMC 2841862. PMID 20190183.
- ↑ Wiesehan K, Willbold D. (2003). "Mirror-image Phage Display: Aiming at the Mirror.". ChemBioChem 4 (9): 811–815. doi:10.1002/cbic.200300570. PMID 12964153.
- ↑ Walensky LD, Kung AL, Escher I et al. (September 2004). "Activation of apoptosis in vivo by a hydrocarbon-stapled BH3 helix". Science 305 (5689): 1466–70. doi:10.1126/science.1099191. PMC 1360987. PMID 15353804.
- ↑ Li L, Thomas RM, Suzuki H, De Brabander JK, Wang X, Harran PG (September 2004). "A small molecule Smac mimic potentiates TRAIL- and TNFalpha-mediated cell death". Science 305 (5689): 1471–4. doi:10.1126/science.1098231. PMID 15353805.
Further reading
- Denicourt C, Dowdy SF (September 2004). "Medicine. Targeting apoptotic pathways in cancer cells". Science 305 (5689): 1411–3. doi:10.1126/science.1102974. PMID 15353788.