Neutrophil granulocyte
| Neutrophil granulocyte | |
|---|---|
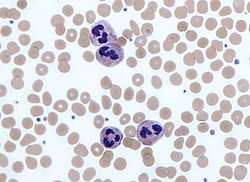 Neutrophils with a segmented nuclei surrounded by erythrocytes, the intra-cellular granules are visible in the cytoplasm (Giemsa stained) | |
| Identifiers | |
| Code | TH H2.00.04.1.02012 |
| Anatomical terminology | |

Neutrophil granulocytes (also known as neutrophils) are the most abundant (40% to 75%) type of white blood cells in mammals and form an essential part of the innate immune system.
They are formed from stem cells in the bone marrow. They are short-lived and highly motile. Neutrophils may be subdivided into segmented neutrophils and banded neutrophils (or bands). They form part of the polymorphonuclear cell family (PMNs) together with basophils and eosinophils.[1][2][3]
The name neutrophil derives from staining characteristics on hematoxylin and eosin (H&E) histological or cytological preparations. Whereas basophilic white blood cells stain dark blue and eosinophilic white blood cells stain bright red, neutrophils stain a neutral pink. Normally, neutrophils contain a nucleus divided into 2–5 lobes.
Neutrophils are a type of phagocyte and are normally found in the bloodstream. During the beginning (acute) phase of inflammation, particularly as a result of bacterial infection, environmental exposure,[4] and some cancers,[5][6] neutrophils are one of the first-responders of inflammatory cells to migrate towards the site of inflammation. They migrate through the blood vessels, then through interstitial tissue, following chemical signals such as Interleukin-8 (IL-8), C5a, fMLP and Leukotriene B4 in a process called chemotaxis. They are the predominant cells in pus, accounting for its whitish/yellowish appearance.
Neutrophils are recruited to the site of injury within minutes following trauma, and are the hallmark of acute inflammation.[7]
Characteristics

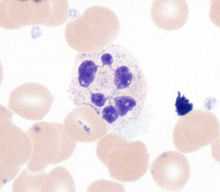
Neutrophil granulocytes have an average diameter of 12-15 micrometers (µm) in peripheral blood smears. When analyzing a pure neutrophil suspension on an automated cell counter, neutrophils have an average diameter of 8–9 µm.
With the eosinophil and the basophil, they form the class of polymorphonuclear cells, named for the nucleus' multilobulated shape (as compared to lymphocytes and monocytes, the other types of white cells). The nucleus has a characteristic lobed appearance, the separate lobes connected by chromatin. The nucleolus disappears as the neutrophil matures, which is something that happens in only a few other types of nucleated cells.[8]:168 In the cytoplasm, the Golgi apparatus is small, mitochondria and ribosomes are sparse, and the rough endoplasmic reticulum is absent.[8]:170 The cytoplasm also contains about 200 granules, of which a third are azurophilic.[8]:170
A minor difference is found between the neutrophils from a male subject and a female subject. The cell nucleus of a neutrophil from a female subject shows a small additional X chromosome structure, known as a "neutrophil drumstick".[8]:174
Neutrophils will show increasing segmentation (many segments of nucleus) as they mature. A normal neutrophil should have 3-5 segments. Hypersegmentation is not normal, and occurs in some disorders, most notably Vitamin B-12 deficiency. This is noted on a manual review of the blood smear, and is positive when most or all or the neutrophils have 5 or more segments.

Neutrophils are the most abundant white blood cells in humans (approximately 1011 are produced daily); they account for approximately 50-70% of all white blood cells (leukocytes). The stated normal range for human blood counts varies between laboratories, but a neutrophil count of 2.5–7.5 x 109/L is a standard normal range. People of African and Middle Eastern descent may have lower counts, which are still normal. A report may divide neutrophils into segmented neutrophils and bands.
When circulating in the bloodstream and unactivated, neutrophils are spherical. Once activated, they change shape and become more amorphous or amoeba-like and can extend pseudopods as they hunt for antigens.[9]
Neutrophils have a preference to engulf refined carbohydrate[10][11][12] (glucose, fructose, sucrose, honey and orange juice[10]) over bacteria.[10] In 1973 Sanchez et al. found that the neutrophil phagocytic capacity to engulf bacteria is affected when simple sugars are digested,[10] and that fasting strengthens the neutrophils' phagocytic capacity to engulf bacteria.[10] However, the digestion of normal starches has no effect. It was concluded that the function, and not the number, of phagocytes in engulfing bacteria was altered by the ingestion of sugars.[10] In 2007 researchers at the Whitehead Institute of Biomedical Research found that even amongst just a selection of sugars, neutrophils can again be picky and engulf preferred sugars.[11][12]
Life span

The average lifespan of (non-activated human) neutrophils in the circulation has been reported by different approaches to be between 5 and 90 hours.[13] Upon activation, they marginate (position themselves adjacent to the blood vessel endothelium), and undergo selectin-dependent capture followed by integrin-dependent adhesion in most cases, after which they migrate into tissues, where they survive for 1–2 days.[14]
Neutrophils are much more numerous than the longer-lived monocyte/macrophage phagocytes. A pathogen (disease-causing microorganism or virus) is likely to first encounter a neutrophil. Some experts hypothesize that the short lifetime of neutrophils is an evolutionary adaptation. The short lifetime of neutrophils minimizes propagation of those pathogens that parasitize phagocytes because the more time such parasites spend outside a host cell, the more likely they will be destroyed by some component of the body's defenses. Also, because neutrophil antimicrobial products can also damage host tissues, their short life limits damage to the host during inflammation.[14]
Neutrophils will often be phagocytosised by macrophages after digestion of pathogens. PECAM-1 and phosphatidylserine on the cell surface are involved in this process.
Chemotaxis
Neutrophils undergo a process called chemotaxis, which allows them to migrate toward sites of infection or inflammation. Cell surface receptors allow neutrophils to detect chemical gradients of molecules such as interleukin-8 (IL-8), interferon gamma (IFN-gamma), C3a, C5a, and Leukotriene B4, which these cells use to direct the path of their migration.
Neutrophils have a variety of specific receptors, including complement receptors, cytokine receptors for interleukins and interferon gamma (IFN-gamma), receptors for chemokines, receptors to detect and adhere to endothelium, receptors for lectins and proteins, and Fc receptors for opsonin.[15]
Anti-microbial function
Being highly motile, neutrophils quickly congregate at a focus of infection, attracted by cytokines expressed by activated endothelium, mast cells, and macrophages. Neutrophils express[16] and release cytokines, which in turn amplify inflammatory reactions by several other cell types.
In addition to recruiting and activating other cells of the immune system, neutrophils play a key role in the front-line defence against invading pathogens. Neutrophils have three methods for directly attacking micro-organisms: phagocytosis (ingestion), release of soluble anti-microbials (including granule proteins), and generation of neutrophil extracellular traps (NETs).[17]
Phagocytosis
Neutrophils are phagocytes, capable of ingesting microorganisms or particles. For targets to be recognised, they must be coated in opsonins—a process known as antibody opsonization.[9] They can internalize and kill many microbes, each phagocytic event resulting in the formation of a phagosome into which reactive oxygen species and hydrolytic enzymes are secreted. The consumption of oxygen during the generation of reactive oxygen species has been termed the "respiratory burst", although unrelated to respiration or energy production.
The respiratory burst involves the activation of the enzyme NADPH oxidase, which produces large quantities of superoxide, a reactive oxygen species. Superoxide decays spontaneously or is broken down via enzymes known as superoxide dismutases (Cu/ZnSOD and MnSOD), to hydrogen peroxide, which is then converted to hypochlorous acid HClO, by the green heme enzyme myeloperoxidase. It is thought that the bactericidal properties of HClO are enough to kill bacteria phagocytosed by the neutrophil, but this may instead be a step necessary for the activation of proteases.[18]
Degranulation
Neutrophils also release an assortment of proteins in three types of granules by a process called degranulation. The contents of these granules have antimicrobial properties, and help combat infection.
| Granule type | Protein |
| azurophilic granules (or "primary granules") | myeloperoxidase, bactericidal/permeability-increasing protein (BPI), defensins, and the serine proteases neutrophil elastase and cathepsin G |
| specific granules (or "secondary granules") | alkaline phosphatase, lysozyme, NADPH oxidase, collagenase, lactoferrin and cathelicidin |
| tertiary granules | cathepsin and gelatinase |
Neutrophil extracellular traps (NETs)
In 2004, Brinkmann and colleagues described a striking observation that activation of neutrophils causes the release of web-like structures of DNA; this represents a third mechanism for killing bacteria.[19] These neutrophil extracellular traps (NETs) comprise a web of fibers composed of chromatin and serine proteases that trap and kill microbes extracellularly. It is suggested that NETs provide a high local concentration of antimicrobial components and bind, disarm, and kill microbes independent of phagocytic uptake. In addition to their possible antimicrobial properties, NETs may serve as a physical barrier that prevents further spread of pathogens. Trapping of bacteria may be a particularly important role for NETs in sepsis, where NETs are formed within blood vessels.[20] Recently, NETs have been shown to play a role in inflammatory diseases, as NETs could be detected in preeclampsia, a pregnancy-related inflammatory disorder in which neutrophils are known to be activated.[21] In addition, NETs are known to exhibit pro-thrombotic effects both in vitro[22] and in vivo.[23][24]
Role in disease
Low neutrophil counts are termed neutropenia. This can be congenital (genetic disorder) or it can develop later, as in the case of aplastic anemia or some kinds of leukemia. It can also be a side-effect of medication, most prominently chemotherapy. Neutropenia makes an individual highly susceptible to infections. Neutropenia can be the result of colonization by intracellular neutrophilic parasites.
In alpha 1-antitrypsin deficiency, the important neutrophil enzyme elastase is not adequately inhibited by alpha 1-antitrypsin, leading to excessive tissue damage in the presence of inflammation – the most prominent one being pulmonary emphysema.
In Familial Mediterranean fever (FMF), a mutation in the pyrin (or marenostrin) gene, which is expressed mainly in neutrophil granulocytes, leads to a constitutively active acute-phase response and causes attacks of fever, arthralgia, peritonitis, and – eventually – amyloidosis.[25]
The Absolute neutrophil count (ANC) is also used in diagnosis and prognosis. The ANC gold standard for determining severity of neutropenia, and thus neutropenic fever. Any ANC < 1500 cells / mm3 is considered neutropenia, but <500 cells / mm3 is considered severe.[26] There is also new research tying ANC to Myocardial infarction as an aid in early diagnosis.[27][28]
Neutrophil antigens
There are five (HNA 1-5) sets of neutrophil antigen recognised.[29] The three HNA-1 antigens (a-c) are located on the low affinity Fc-γ receptor IIIb (FCGR3B :CD16b) The single known HNA-2a antigen is located on CD177. The HNA-3 antigen system has two antigens (3a and 3b) which are located on the seventh exon of the CLT2 gene (SLC44A2). The HNA-4 and HNA-5 antigen systems each have two known antigens (a and b) and are located in the β2 integrin. HNA-4 is located on the αM chain (CD11b) and HNA-5 is located on the αL integrin unit (CD11a).
Video
-
A rapidly moving neutrophil can be seen taking up several conidia over an imaging time of 2 hours with one frame every 30 seconds.
-
A neutrophil can be seen here selectively taking up several Candida yeasts (fluorescently labeled in green) despite several contacts with Aspergillus fumigatus conidia (unlabeled, white/clear) in a 3-D collagen matrix. Imaging time was 2 hours with one frame every 30 seconds.
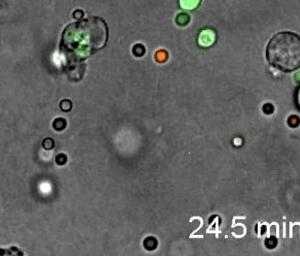
Neutrophils display highly directional amoeboid motility in infected footpad and phalanges. Intravital imaging was performed in the footpad path of LysM-eGFP mice 20 minutes after infection with Listeria monocytogenes.[30]
Additional images
-
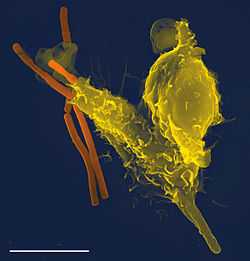
A scanning electron microscope image of a single neutrophil (yellow), engulfing anthrax bacteria (orange)
-

Blood cell lineage
-
_diagram.png)
More complete lineages (very large)
References
- ↑ Witko-Sarsat, V; Rieu P, Descamps-Latscha B, Lesavre P, Halbwachs-Mecarelli L (2000). "Neutrophils: molecules, functions and pathophysiological aspects". Lab Invest 80 (5): 617–53. doi:10.1038/labinvest.3780067. PMID 10830774.
- ↑ Klebanoff, SJ; Clark, RA (1978). "The Neutrophil: Function and Clinical Disorders". Elsevier/North-Holland Amsterdam. ISBN 0-444-80020-4.
- ↑ Nathan, C (Mar 2006). "Neutrophils and immunity: challenges and opportunities". Nature Reviews Immunology 6 (March): 173–82. doi:10.1038/nri1785. ISSN 1474-1733. PMID 16498448.
- ↑ Jacobs, L; Nawrot, Tim S; De Geus, Bas; Meeusen, Romain; Degraeuwe, Bart; Bernard, Alfred; Sughis, Muhammad; Nemery, Benoit; Panis, Luc (Oct 2010). "Subclinical responses in healthy cyclists briefly exposed to traffic-related air pollution". Environmental Health 9 (64): 64. doi:10.1186/1476-069X-9-64. PMC 2984475. PMID 20973949.
- ↑ Waugh, DJ; Wilson, C. (Nov 2008). "The interleukin-8 pathway in cancer". Clinical Cancer Research 14 (21): 6735–41. doi:10.1158/1078-0432.CCR-07-4843. ISSN 1078-0432. PMID 18980965.
- ↑ De Larco, JE; Wuertz, BR; Furcht, LT (Aug 2004). "The Potential Role of Neutrophils in Promoting the Metastatic Phenotype of Tumors Releasing Interleukin-8". Clinical Cancer Research 10 (15): 4895–900. doi:10.1158/1078-0432.CCR-03-0760. ISSN 1078-0432. PMID 15297389.
- ↑ Cohen, Stephen; Burns, Richard C. (2002). Pathways of the Pulp (8th ed.). St. Louis: Mosby. p. 465.
- ↑ 8.0 8.1 8.2 8.3 Zucker-Franklin, Dorothy; Greaves, M. F.; Grossi, C. E.; Marmont, A. M. (1988). "Neutrophils". Atlas of Blood Cells: Function and Pathology 1 (2nd ed.). Philadelphia: Lea & Ferbiger. ISBN 0-8121-1094-3.
- ↑ 9.0 9.1 Edwards, Steven W. (1994). Biochemistry and physiology of the neutrophil. Cambridge University Press. p. 6. ISBN 0-521-41698-1.
- ↑ 10.0 10.1 10.2 10.3 10.4 10.5 Albert Sanchez, J. L. Reeser, H. S. Lau, P. Y. Yahiku, R. E. Willard, P. J. McMillan, S. Y. Cho, A. R. Magie, and U. D. Register (1973). "Role of sugars in human neutrophilic phagocytosis". The American Society for Clinical Nutrition. Retrieved 2013-09-08.
These data suggest that the function and not the number of phagocytes was altered by ingestion of sugars. This implicates glucose and other simple carbohydrates in the control of phagocytosis and shows that the effects last for at least 5 hr. On the other hand, a fast of 36 or 60 hr significantly increased (P < 0.001) the phagocytic index.
- ↑ 11.0 11.1 Rubin-Bejerano, I.; Abeijon, C.; Magnelli, P.; Grisafi, P.; Fink, G. R. (July 2007). "Phagocytosis by human neutrophils is stimulated by a unique fungal cell wall component". Cell Host Microbe 2 (1): 55–67. doi:10.1016/j.chom.2007.06.002. PMC 2083279. PMID 18005717.
- ↑ 12.0 12.1 Kneller, Alyssa (2007). "White blood cells are picky about sugar". Whitehead Institute. Retrieved 2013-08-09.
- ↑ Tak T, Tesselaar K, Pillay J, Borghans JA, Koenderman L (2013). "What's your age again? Determination of human neutrophil half-lives revisited". Journal of Leukocyte Biology 94 (4): 595–601. doi:10.1189/jlb.1112571. PMID 23625199.
- ↑ 14.0 14.1 Wheater, Paul R.; Stevens, Alan (2002). Wheater's basic histopathology: a colour atlas and text (PDF). Edinburgh: Churchill Livingstone. ISBN 0-443-07001-6.
- ↑ Serhan, Charles N.; Ward, Peter A.; Gilroy, Derek W. (2010). Fundamentals of Inflammation. Cambridge University Press. pp. 53–54. ISBN 0-521-88729-1.
- ↑ Ear, T; McDonald, PP (2008). "Cytokine generation, promoter activation, and oxidant-independent NF-kappaB activation in a transfectable human neutrophilic cellular model". BMC Immunol. 9: 14. doi:10.1186/1471-2172-9-14. PMC 2322942. PMID 18405381.
- ↑ Hickey, MJ; Kubes, P (2009). "Intravascular immunity: the host–pathogen encounter in blood vessels". Nature Reviews Immunology (Nature Publishing Group) 9 (5): 364–75. doi:10.1038/nri2532. PMID 19390567.
- ↑ Segal, AW (2005). "How neutrophils kill microbes". Annu Rev Immunol 9 (5): 197–223. doi:10.1146/annurev.immunol.23.021704.115653. PMC 2092448. PMID 15771570.
- ↑ Brinkmann, Volker; Ulrike Reichard, Christian Goosmann, Beatrix Fauler, Yvonne Uhlemann, David S. Weiss, Yvette Weinrauch, Arturo Zychlinsky (5 March 2004). "Neutrophil Extracellular Traps Kill Bacteria". Science (AAAS) 303 (5663): 1532–1535. doi:10.1126/science.1092385. ISSN 0036-8075. PMID 15001782. Retrieved 2007-04-09.
- ↑ Clark SR, Ma AC, Tavener AS, McDonald B, Goodarzi Z, Kelly MM, Patel KD, Chakrabarti S, McAvoy E, Sinclair GD, Keys EM, Allen-Vercoe E, DeVinney R, Doig CJ, Green FHY and Kubes P (Apr 2007). "Platelet Toll-Like Receptor-4 Activates Neutrophil Extracellular Traps to Ensnare Bacteria in Endotoxemic and Septic Blood" (PDF). Nature Medicine (Nature Publishing Group) 13 (4): 463–9. doi:10.1038/nm1565. ISSN 1078-8956. PMID 17384648.
- ↑ Gupta, AK; Hasler, P; Holzgreve, W; Hahn, S (Jun 2007). "Neutrophil NETs: a novel contributor to preeclampsia-associated placental hypoxia?". Semin Immunopathol 29 (2): 163–7. doi:10.1007/s00281-007-0073-4. ISSN 1863-2297. PMID 17621701.
- ↑ Fuchs, TA; Brill, A, Duerschmied, D, Schatzberg, D, Monestier, M, Myers DD, Jr, Wrobleski, SK, Wakefield, TW, Hartwig, JH, Wagner, DD (Sep 7, 2010). "Extracellular DNA traps promote thrombosis". Proceedings of the National Academy of Sciences of the United States of America 107 (36): 15880–5. doi:10.1073/pnas.1005743107. PMC 2936604. PMID 20798043.
- ↑ Brill, A; Fuchs, TA, Savchenko, A, Thomas, GM, Martinod, K, De Meyer, SF, Bhandari, AA, Wagner, DD (Nov 1, 2011). "Neutrophil Extracellular Traps Promote Deep Vein Thrombosis in Mice". Journal of thrombosis and haemostasis : JTH 10 (1): 136–44. doi:10.1111/j.1538-7836.2011.04544.x. PMC 3319651. PMID 22044575.
- ↑ Borissoff, JI; ten Cate, H (September 2011). "From neutrophil extracellular traps release to thrombosis: an overshooting host-defense mechanism?". Journal of thrombosis and haemostasis : JTH 9 (9): 1791–4. doi:10.1111/j.1538-7836.2011.04425.x. PMID 21718435.
- ↑ Ozen, S (Jul 2004). "Familial mediterranean fever: revisiting an ancient disease". European Journal of Pediatrics 162 (7–8): 449–54. doi:10.1007/s00431-003-1223-x. ISSN 0340-6199. PMID 12751000.
- ↑ Al-Gwaiz, LA; Babay, HH (2007;16(5):344-7.). "The diagnostic value of absolute neutrophil count, band count and morphologic changes of neutrophils in predicting bacterial infections.". Med Princ Pract. 16 (5): 344–7. doi:10.1159/000104806. PMID 17709921. Check date values in:
|date=(help) - ↑ Khan, HA et al. (2012). "Blood cell counts and their correlation with creatine kinase and C-reactive protein in patients with acute myocardial infarction.". Int J Clin Exp Med. 5 (1): 50–5. PMID 22328948.
- ↑ Basili, S et al. (2004). "Absolute neutrophil counts and fibrinogen levels as an aid in the early diagnosis of acute myocardial infarction.". Acta Cardiol 59 (2): 135–40. doi:10.2143/ac.59.2.2005167. PMID 15139653.
- ↑ Chu HT, Lin H, Tsao TT, Chang CF, Hsiao WW, Yeh TJ, Chang CM, Liu YW, Wang TY, Yang KC, Chen TJ, Chen JC, Chen KC, Kao CY (2013). "Genotyping of human neutrophil antigens (HNA) from whole genome sequencing data". BMC Med Genomics 6 (1): 31. doi:10.1186/1755-8794-6-31. PMC 3849977. PMID 24028078.
- ↑ Graham D.B., Zinselmeyer B.H., Mascarenhas F., Delgado R., Miller M.J., Swat W.; Zinselmeyer; Mascarenhas; Delgado; Miller; Swat (2009). Unutmaz, Derya, ed. "ITAM signaling by Vav family Rho guanine nucleotide exchange factors regulates interstitial transit rates of neutrophils in vivo". PLoS ONE 4 (2): e4652. doi:10.1371/journal.pone.0004652. PMC 2645696. PMID 19247495.
External Links
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||