Nest (protein structural motif)
The Nest is a type of protein structural motif. Peptide nests are small anion-binding molecular features of proteins and peptides.[1][2][3][4][5][6][7] Each consists of the main chain atoms only of three consecutive amino acid residues. The main chain NH groups bind the anions while the side chain atoms are often not involved. Proline residues lack NH groups so are rare in nests. About one in 12 of amino acid residues in proteins, on average, belongs to a nest. Two websites are available for examining nests in proteins, Motivated Proteins: ;[8] or PDBeMotif: .[9]
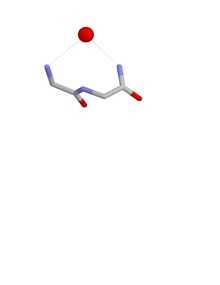
Nest conformations
The conformation of a nest is such that one of more of the NH groups of the first, second and third amino acid residues are liable to be hydrogen bonded to a negatively charged, or partially negatively charged, atom, often an oxygen atom. These main chain atoms form a concavity called a nest into which an anionic atom fits. Such anionic atoms are sometimes called eggs and more than one egg may occur bound to a nest. The oxyanion hole of the intestinal serine proteases is a functional example of a nest. Another occurs in the antibiotic peptide vancomycin which binds a key carboxylate group needed during bacterial cell wall synthesis, thereby preventing the cells from multiplying.
Nests are defined by the conformation of the main chain atoms, namely the phi,psi dihedral angles of the first two amino acids in the nest. Nests vary in their degree of concavity. A few have so little that the concavity is lost. Such peptides are less liable to act as anion binding sites. The specificity filter of the potassium channel[10] and the channel of aquaporin exhibit this more linear conformation in which the carbonyl groups are employed by proteins to transport molecules across membranes. This near-linear conformation is also that found in a strand of alpha sheet[11][12][13]
Compound nests
If two nests overlap such that residue i+1 of the first nest is residue i of the second nest, a compound nest is formed. This has four NH groups instead of three. If three nests overlap such that residues i+1 and i+2 of the first nest are both residue i of a nest, a wider compound nest is formed with five NH groups, and so on. The main chain atoms form part of a ring with the NH groups all pointing roughly inwards, towards the centre of the ring. Because their concavities are often wider than simple nests, compound nests are commonly employed by proteins for binding anions such as phosphates, as in the P-loop or Walker motifs, and in iron-sulphur clusters.
Types of nest
Simple nests are of two kinds called RL and LR depending on the sign of the phi angles of the first two nest residues. R residues have negative phi values (as in right-handed alpha-helices) and L residues have positive phi values (as in the left-handed alpha helix). Eighty percent of nests are RL and 20% are LR. When two nests overlap they may be RLR or LRL. When three nests overlap they may be RLRL or LRLR, and so on.
Every Schellman loop incorporates an RL nest in the last three of its six residues. The nest binds carbonyl oxygen atoms preceding it in sequence.
A number of antibody proteins have RLR nests within the hairpin loops of their H-chain CDRs (complementarity determining regions), with the capacity to bind the phosphates of phosphorylated serines and threonines.[14]
References
- ↑ Watson, JD; Milner-White (2002). "A novel main-chain anion-binding site in proteins: The nest. A particular combination of phi,psi values in successive residues gives rise to anion-binding sites that occur commonly and are found often at functionally important regions". Journal of Molecular Biology 315 (2): 171–182. doi:10.1006/jmbi.2001.5227. PMID 11779237.
- ↑ Pal, D; Suhnel (2002). "New principles of protein structure: nests, eggs and what next?". Angewandte Chemie Int Ed 41 (24): 4663–4665. doi:10.1002/anie.200290009.
- ↑ Milner-White, EJ; Nissink (2004). Acta Crystallographica Section D D60: 1935–1942. Missing or empty
|title=(help) - ↑ Pajewski, R; Ferdani (2005). "Cation Dependence of Chloride Ion Complexation by Open-Chained Receptor Molecules in Chloroform Solution". Journal of the American Chemical Society 127 (51): 18281–18295. doi:10.1021/ja0558894.
- ↑ Berkessel, A; Koch (2006). "Asymmetric enone epoxidation by solid-phase bound peptides: further evidence for catalyst helicity and catalytic activity of individual strands". Biopolymers 84 (1): 90–96. doi:10.1002/bip.20413.
- ↑ Milner-White, EJ; Russell (2006). "Predicting the conformations of proteins and peptides in early evolution". Biology Direct 3: 3.
- ↑ Watson, JD; Laskowski (2005). "ProFunc: a server for predicting protein function from 3D structure". Nucleic Acids Research 33 (Web Server): W89–W93. doi:10.1093/nar/gki414.
- ↑ Leader, DP; Milner-White (2009). "Motivated Proteins: A web application for studying small three-dimensional protein motifs". BMC Bioinformatics 10 (1): 60. doi:10.1186/1471-2105-10-60. PMC 2651126. PMID 19210785.
- ↑ Golovin, A; Henrick (2008). "MSDmotif: exploring protein sites and motifs". BMC Bioinformatics 9 (1): 312. doi:10.1186/1471-2105-9-312. PMC 2491636. PMID 18637174.
- ↑ Watson, JD; Milner-White (2002). "The conformations of polypeptide chains where the main-chain parts of successive residues are enantiomeric. Their occurrence in cation and anion-binding regions of proteins". Journal of Molecular Biology 315 (15): 183–191. doi:10.1006/jmbi.2001.5228. PMID 11779238.
- ↑ Milner-White, EJ; Watson (2006). "Amyloid formation may involve alpha- to beta sheet interconversion via peptide plane flipping". Structure 14 (9): 1369–1376. doi:10.1016/j.str.2006.06.016.
- ↑ Hayward, S; Milner-White (2008). "The geometry of α-sheet: Implications for its possible function as amyloid precursor in proteins". Proteins 71 (1): 415–425. doi:10.1002/prot.21717.
- ↑ Hayward, S; Milner-White (2011). "Simulation of the β- to α-sheet transition results in a twisted sheet for antiparallel and an α-nanotube for parallel strands: implications for amyloid formation". Proteins 79 (11): 3193–3207. doi:10.1002/prot.23154.
- ↑ Koerber, JT; Thomsen ND (2014). "Nature-inspired design of motif-specific antibody scaffolds". Nature Biotechnology 31 (10): 916–921. doi:10.1038/nbt.2672.