Heterochromatin
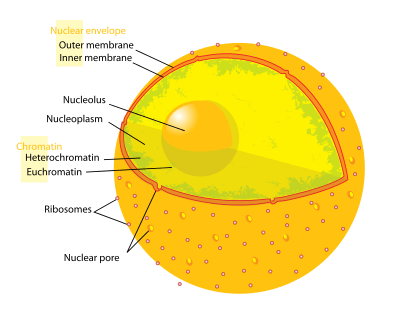
Heterochromatin is a tightly packed form of DNA, which comes in different varieties. These varieties lie on a continuum between the two extremes of constitutive and facultative heterochromatin. Both play a role in the expression of genes, where constitutive heterochromatin can affect the genes near them (position-effect variegation) and where facultative heterochromatin is the result of genes that are silenced through a mechanism such as histone deacetylation or piRNA through RNAi. Constitutive heterochromatin is usually repetitive and forms structural functions such as centromeres or telomeres, in addition to acting as an attractor for other gene-expression or repression signals. Facultative heterochromatin is not repetitive and although it shares the compact structure of constitutive heterochromatin, facultative heterochromatin can, under specific developmental or environmental signaling cues, lose its condensed structure and become transcriptionally active.[1] Heterochromatin has been associated with the di and tri-methylation of H3K9 in certain portions of the genome.[2]
Structure
Chromatin is found in two varieties: euchromatin and heterochromatin.[3] Originally, the two forms were distinguished cytologically by how intensely they stained - the euchromatin is less intense, while heterochromatin stains intensely, indicating tighter packing. Heterochromatin is usually localized to the periphery of the nucleus. Despite this early dichotomy, recent evidence in both animals [4] and plants [5] has suggested that there are more than two distinct heterochromatin states, and it may in fact exist in four or five 'states', each marked by different combinations of epigenetic marks.
Heterochromatin mainly consists of genetically inactive satellite sequences,[6] and many genes are repressed to various extents, although some cannot be expressed in euchromatin at all.[7] Both centromeres and telomeres are heterochromatic, as is the Barr body of the second, inactivated X-chromosome in a female.
Function
Heterochromatin has been associated with several functions, from gene regulation to the protection of the integrity of chromosomes;[8] some of these roles can be attributed to the dense packing of DNA, which makes it less accessible to protein factors that usually bind DNA or its associated factors. For example, naked double-stranded DNA ends would usually be interpreted by the cell as damaged or viral DNA, triggering cell cycle arrest, DNA repair, or destruction of the DNA fragment such as by endonucleases in bacteria.
Some regions of chromatin are very densely packed with fibers displaying a condition comparable to that of the chromosome at mitosis. Heterochromatin is generally clonally inherited; when a cell divides the two daughter cells will typically contain heterochromatin within the same regions of DNA, resulting in epigenetic inheritance. Variations cause heterochromatin to encroach on adjacent genes or recede from genes at the extremes of domains. Transcribable material may be repressed by being positioned (in cis) at these boundary domains. This gives rise to different levels of expression from cell to cell,[9] which may be demonstrated by position-effect variegation.[10] Insulator sequences may act as a barrier in rare cases where constitutive heterochromatin and highly active genes are juxtaposed (e.g. the 5'HS4 insulator upstream of the chicken β-globin locus,[11] and loci in two Saccharomyces spp.[12][13]).
Constitutive heterochromatin
All cells of a given species that will package the same regions of DNA in constitutive heterochromatin, and thus in all cells any genes contained within the constitutive heterochromatin will be poorly expressed. For example, all human chromosomes 1, 9, 16, and the Y-chromosome contain large regions of constitutive heterochromatin. In most organisms, constitutive heterochromatin occurs around the chromosome centromere and near telomeres.
Facultative heterochromatin
The regions of DNA packaged in facultative heterochromatin will not be consistent between the cell types within a species, and thus a sequence in one cell that is packaged in facultative heterochromatin (and the genes within are poorly expressed) may be packaged in euchromatin in another cell (and the genes within are no longer silenced). However, the formation of facultative heterochromatin is regulated, and is often associated with morphogenesis or differentiation. An example of facultative heterochromatin is X-chromosome inactivation in female mammals: one X chromosome is packaged as facultative heterochromatin and silenced, while the other X chromosome is packaged as euchromatin and expressed.
Among the molecular components that appear to regulate the spreading of heterochromatin include the Polycomb-group proteins and non-coding genes such as Xist. The mechanism for such spreading is still a matter of controversy.[14]
Yeast heterochromatin
Saccharomyces cerevisiae, or budding yeast, is a model eukaryote and its heterochromatin has been defined thoroughly. Although most of its genome can be characterized as euchromatin, S. cerevisiae has regions of DNA that are transcribed very poorly. These loci are the so-called silent mating type loci (HML and HMR), the rDNA (encoding ribosomal RNA), and the sub-telomeric regions. Fission yeast (Schizosaccharomyces pombe) uses another mechanism for heterochromatin formation at its centromeres. Gene silencing at this location depends on components of the RNAi pathway. Double-stranded RNA is believed to result in silencing of the region through a series of steps.
In the fission yeast Schizosaccharomyces pombe two RNAi complexes, the RNAi-induced transcriptional gene silencing (RITS) complex and the RNA-directed RNA polymerase complex (RDRC), are part of an RNAi machinery involved in the initiation, propagation and maintenance of heterochromatin assembly. These two complexes localize in a siRNA-dependent manner on chromosomes, at the site of heterochromatin assembly. RNA polymerase II synthesizes a transcript that serves as a platform to recruit RITS, RDRC and possibly other complexes required for heterochromatin assembly.[15][16] Both RNAi and an exosome-dependent RNA degradation process contribute to heterochromatic gene silencing. These mechanisms of Schizosaccharomyces pombe may occur in other eukaryotes.[17] A large RNA structure called RevCen has also been implicated in the production of siRNAs to mediate heterochromatin formation in some fission yeast.[18]
See also
- Centric Heterochromatin
External links
- Histology image: 20102loa — Histology Learning System at Boston University
References
- ↑ Oberdoerffer, P; Sinclair, D (2007). "The role of nuclear architecture in genomic instability and ageing". Nature Reviews Molecular Cell Biology 8: 692–702. doi:10.1038/nrm2238.
- ↑ Rosenfeld, Jeffrey A; Wang, Zhibin; Schones, Dustin; Zhao, Keji; Desalle, Rob; Zhang, Michael Q (31 March 2009). "Determination of enriched histone modifications in non-genic portions of the human genome". BMC Genomics 10 (1): 143. doi:10.1186/1471-2164-10-143. PMC 2667539. PMID 19335899.
- ↑ Elgin, S.C. (1996). "Heterochromatin and gene regulation in Drosophila". Current Opinion in Genetics & Development 6 (2): 193–202. doi:10.1016/S0959-437X(96)80050-5. ISSN 0959-437X.
- ↑ van Steensel, B. (2011). "Chromatin: constructing the big picture". The EMBO Journal 30 (10): 1885–95. doi:10.1038/emboj.2011.135. PMC 3098493. PMID 21527910.
- ↑ Roudier, François "et al." (2011). "Integrative epigenomic mapping defines four main chromatin states in Arabidopsis". The EMBO Journal 30 (10): 1928–1938. doi:10.1038/emboj.2011.103. PMC 3098477. PMID 21487388.
|first2=missing|last2=in Authors list (help);|first3=missing|last3=in Authors list (help);|first4=missing|last4=in Authors list (help);|first5=missing|last5=in Authors list (help);|first6=missing|last6=in Authors list (help);|first7=missing|last7=in Authors list (help);|first8=missing|last8=in Authors list (help);|first9=missing|last9=in Authors list (help);|first10=missing|last10=in Authors list (help);|first11=missing|last11=in Authors list (help);|first12=missing|last12=in Authors list (help);|first13=missing|last13=in Authors list (help);|first14=missing|last14=in Authors list (help);|first15=missing|last15=in Authors list (help);|first16=missing|last16=in Authors list (help);|first17=missing|last17=in Authors list (help);|first18=missing|last18=in Authors list (help);|first19=missing|last19=in Authors list (help);|first20=missing|last20=in Authors list (help);|first21=missing|last21=in Authors list (help);|first22=missing|last22=in Authors list (help);|first23=missing|last23=in Authors list (help) - ↑ Lohe, A.R. et al. (August 1, 1993). "Mapping Simple Repeated DNA Sequences in Heterochromatin of Drosophila Melanogaster". Genetics 134 (4): 1149–74. ISSN 0016-6731. PMC 1205583. PMID 8375654.
- ↑ Lu, B.Y. et al. (June 1, 2000). "Heterochromatin protein 1 is required for the normal expression of two heterochromatin genes in Drosophila". Genetics 155 (2): 699–708. ISSN 0016-6731. PMC 1461102. PMID 10835392.
- ↑ Grewal SIS & Jia S (January 2007). "Heterochromatin revisited". Nature Reviews Genetics 8 (1): 35–. doi:10.1038/nrg2008. PMID 17173056. Retrieved 18 September 2013.
An up-to-date account of the current understanding of repetitive DNA, which usually doesn't contain genetic information. If evolution makes sense only in the context of the regulatory control of genes, we propose that heterochromatin, which is the main form of chromatin in higher eukaryotes, is positioned to be a deeply effective target for evolutionary change. Future investigations into assembly, maintenance and the many other functions of heterochromatin will shed light on the processes of gene and chromosome regulation.
- ↑ Fisher, Amanda G.; Matthias Merkenschlager (April 2002). "Gene silencing, cell fate and nuclear organisation". Current Opinion in Genetics & Development 12 (2): 193–7. doi:10.1016/S0959-437X(02)00286-1. ISSN 0959-437X. PMID 11893493.
- ↑ Zhimulev, I.F. et al. (December 1986). "Cytogenetic and molecular aspects of position effect variegation in Drosophila melanogaster". Chromosoma 94 (6): 492–504. doi:10.1007/BF00292759. ISSN 1432-0886.
- ↑ Burgess-Beusse, B et al. (December 2002). "The insulation of genes from external enhancers and silencing chromatin". Proc. Natl Acad. Sci. USA 9 (Suppl 4): 16433–7. doi:10.1073/pnas.162342499. PMC 139905. PMID 12154228.
- ↑ Noma, K. et al. (August 2001). "transitions in distinct histone H3 methylation patterns at the heterochromatin domain boundaries". Science 293 (5532): 1150–5. doi:10.1126/science.1064150. PMID 11498594.
- ↑ Donze, D. & R.T. Kamakaka (2000). "RNA polymerase III and RNA polymerase II promoter complexes are heterochromatin barriers in Saccharomyces cerevisiae". The EMBO Journal 20 (3): 520–31. doi:10.1093/emboj/20.3.520. PMC 133458. PMID 11157758.
- ↑ Talbert PB, Henikoff S (October 2006). "Spreading of silent chromatin: inaction at a distance". Nature Reviews Genetics 7 (10): 793–803. doi:10.1038/nrg1920. PMID 16983375.
- ↑ Kato, H. et al. (2005). Science 309: 467–469. doi:10.1126/science.1114955. Missing or empty
|title=(help) - ↑ Djupedal, I. et al. (2005). Genes & Development 19: 2301–2306. doi:10.1101/gad.344205. Missing or empty
|title=(help) - ↑ Vavasseur et al. (2008). "Heterochromatin Assembly and Transcriptional Gene Silencing under the Control of Nuclear RNAi: Lessons from Fission Yeast". RNA and the Regulation of Gene Expression: A Hidden Layer of Complexity. Caister Academic Press. ISBN 978-1-904455-25-7.
- ↑ Djupedal I, Kos-Braun IC, Mosher RA et al. (December 2009). "Analysis of small RNA in fission yeast; centromeric siRNAs are potentially generated through a structured RNA". EMBO J. 28 (24): 3832–44. doi:10.1038/emboj.2009.351. PMC 2797062. PMID 19942857.
- Avramova Z (May 2002). "Heterochromatin in Animals and Plants. Similarities and Differences". Plant Physiology 129 (1): 40–9. doi:10.1104/pp.010981. PMC 1540225. PMID 12011336.
- Caron H et al. (2001). "The Human Transcriptome Map: Clustering of Highly Expressed Genes in Chromosomal Domains". Science 291 (5507): 1289–92. doi:10.1126/science.1056794. PMID 11181992.
|first2=missing|last2=in Authors list (help);|first3=missing|last3=in Authors list (help);|first4=missing|last4=in Authors list (help);|first5=missing|last5=in Authors list (help);|first6=missing|last6=in Authors list (help);|first7=missing|last7=in Authors list (help);|first8=missing|last8=in Authors list (help);|first9=missing|last9=in Authors list (help);|first10=missing|last10=in Authors list (help);|first11=missing|last11=in Authors list (help);|first12=missing|last12=in Authors list (help);|first13=missing|last13=in Authors list (help)
| ||||||||||||||||||||||||||||||||||||||||||||||