HLA-DQ2
| HLA-DQ2 (MHC Class II, DQ) | |||
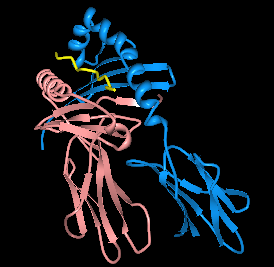 | |||
| Illustration of HLA-DQ2.5 with deamidated gliadin peptide in the binding pocket.1s9v | |||
| Cis-haplotype | Haplotype | ||
| isoform, | subtype | DQA1 | DQB1 |
| DQ α2β2 | DQ2.2 | *0201 | *0202 |
| DQ α3β2 | DQ2.3 | *0303 | *0202 |
| DQ α5β2 | DQ2.5 | *0501 | *0201 |
HLA-DQ2 (DQ2) is a serotype group within HLA-DQ (DQ) serotyping system. The serotype is determined by the antibody recognition of β2 subset of DQ β-chains. The β-chain of DQ is encoded by HLA-DQB1 locus and DQ2 are encoded by the HLA-DQB1*02 allele group. This group currently contains two common alleles, DQB1*0201 and DQB1*0202. HLA-DQ2 and HLA-DQB1*02 are almost synonymous in meaning. DQ2 β-chains combine with α-chains, encoded by genetically linked HLA-DQA1 alleles, to form the cis-haplotype isoforms. These isoforms, nicknamed DQ2.2 and DQ2.5, are also encoded by the DQA1*0201 and DQA1*0501 genes, respectively.
DQ2 is most common in Western Europe, North Africa and East Africa. Highest frequencies are observed in parts of Spain and Ireland; this distribution correlates with the frequency of two of the most prevalent autoimmune diseases. There is also an increase in DQB1*0201 in Central Asia, peaking in Kazakhstan and declining slowly east to west into China and finally Southeast Asia. DQA1*0501 : DQB1*0201. DQ2.5 is one of the most predisposing factors for autoimmune disease. DQ2.5 is encoded, often, by a haplotype associated with a large number of diseases. This haplotype, HLA A1-B8-DR3-DQ2, is associated with diseases in which HLA-DQ2 has suspect involvement. Direct involvement of DQ2 is certain in coeliac disease.
Serology
| DQB1* | DQ2 | Sample |
| allele | % | size (N) |
| *0201 | 98 | 9175 |
| *0202 | 87 | 656 |
| DQ | DQ | DQ | Freq | |||||
|---|---|---|---|---|---|---|---|---|
| Serotype | cis-isoform | Subtype | A1 | B1 | %[2] | rank | ||
| DQ2 | α5-β2 | 2.5 | 0501♣ | 0201 | 13. | 16 | 2nd | |
| α2-β2 | 2.2 | 0201 | 0202 | 11. | 08 | 3rd | ||
| α3-β2 | 2.3 | 0302♠ | 0202 | 0. | 08 | |||
| ♠DQA1*0302 & *0303 not reolved; ♣DQB1*0501 & *0505 , and some *0303 are resolvable by haplotype | ||||||||
Serotyping efficiency. The serotyping efficiency of HLA-DQ2 is among the highest of the antisera. DQ2 antibodies can be used to effectively type DQ2 bearing individuals, however antibody may detect DQB1*0303.
Alleles
DQB1*0201
The DQB1*0201 allele is genetically linked to DQA1*0501 and DRB1*03. With DQA1*0501 it forms the DQ2.5cis encoding haplotype, with DRB1*03 it becomes part of the DR3-DQ2 (DR-DQ) serologically defined haplotype. With DQA1*0501 the allele is most frequently found in coeliac disease. With DR3 this DQ2 has the second strongest linkage to Type 1 diabetes, and when paired with HLA-DQ8 is the most abundant phenotype found in late onset, "Type1-Type2" diabetes mellitus type 1. As the incidence of coeliac disease is about 1% this allele is associated with more autoimmune disease relative to any other DQ haplotype.
There are ambiguities regarding DQB1*0201 in the literature, some low resolution typing kits detect *0202 as *0201 and are presented as *0201 in the literature without distinction. DQB1*0201 in Europe is frequently found in the HLA A1-B8-DR3-DQ2 haplotype.
DQB1*0202
This allele is linked to several DQA1* alleles, the linkage with DQA1*0201 forms the DQ2.2 haplotype and linkage with DQA1*0303 forms the DQ2.3 haplotype. In Africa the DQA1*0501 haplotype is also linked rarely to DQB1*0202 and may represent that ancestral form of the DQ2.5 haplotype.
DQ2 isoforms and DQ2 haplotypes
|




DQ2.5
DQ2.5 refers to either a protein isoform and a genetic (chromosomal) haplotype. DQ2.5 isoform or heterodimer is shorthand for the cell surface receptor HLA-DQ α5β2. Frequently called 'the DQ2 heterodimer', the DQ2.5 isoform is actually one of two common DQ heterodimers, the other being DQ2.2. DQ2.5 haplotype is created by close genetic linkage of two alleles, written as a haplotype, DQA1*0501:DQB1*0201. The haplotype encodes DQ2.5cis isoform, referring to the cis arrangement of the DQA1*0501 and DQB1*0201 on the same variant of chromosome 6. The isoform can also be encoded trans-haplotype (between two sister chromosomes) forming the DQ2.5trans isoform. This isoform occurs when a person has the DQ7.5/DQ2.2 phenotype.
DQ2.5 and the linked DR3 are associated with probably the greatest frequency of autoimmune occurrence relative to any other haplotypes. The haplotype is positively associated with coeliac disease, dermatitis herpetiformis, juvenile diabetes, Lambert-Eaton myasthenic syndrome (LEMS), Sjögren's syndrome, and autoimmune hepatitis (although significant proportion of the risk is secondary to coeliac disease). DR3 and/or DQ2.5 are linked to the following diseases: Moreen's ulceration,[5] "bout onset" multiple sclerosis,[6] Grave's disease[7] and systemic lupus erythematosus.[8]
DQ2.2
DQ2.2 is shorthand for the DQ α2β2 heterodimeric isoform. The isoform is encoded almost exclusively by the DQA1*0201:DQB1*0202 haplotype. The haplotype is linked to DR7. A small percentage of coeliac disease are associated with this haplotype, and some disease causing gliadins are presented by DQ2.2. The haplotype is found at high frequencies in the Mediterranean and West Africa. The Eurasian geographic distribution of DQ2.2 is slightly greater than DQ2.5. Compared to DQ2.5, the freqeuncy in Sardinia is low, but in Iberia it is high reaching a maximum frequency of ~30% in Northern Iberia, and half that in the British Ilses. It extends along the Mediterranean and Africa at relatively high frequency and is found in high frequencies in some Central Asian, Mongolians, and Han Chinese. It does not appear to have an indigenous presence in the West Pacific Rim or the New World and DQ2.2 presence in Southeast Asia and Indonesia is likely the result of gene flow from India and China in post-neolithic times. The haplotype shows considerable diversity in Africa and this has translated to Iberia with 2 addition haplotypes, DQA1*0303:DQB1*0202 and DR7:DQA1*0201:DQB1*0303. The expansion of DQ2.2 into Europe appears to have been slightly later or biased by some constriction between Iberia and the rest of the continent.
DQ2.3
DQ2.3 is shorthand for the DQ α3β2 heterodimeric isoform. The isoform is encoded by DQA1*0303:DQB1*0202 haplotype. The isoform can also be produced by phenotypes were one haplotype is DQ4.3, DQ7.3, DQ8.1, DQ9.3 and the other haplotype is DQ2.2 or DQ2.5. Therefore the haplotype encoded receptor is a DQ2.3cis isoform which is genetically linked to DR7 By serology DR7-DQ2 cannot discriminate DQ2.2 from DQ2.3 haplotypes, and therefore DQA1 typing is required.
Other isoforms
DQ2 beta chains can pair with trans to other alpha chain. However there is no precedance in cis isoforms for DQ2,4,7,8, or 9 binding DQ1 (DQA1*01) alpha chains. DQA1*03, *05 chains process to near identical alpha chains. The *04 chain can potentially complex with DQ2 forming DQ2.4. In East Asia there is the possibility of DQ2.6 resulting from pairing with DQA1*0601.
In celiac disease
DQ2 represents the second highest risk factor for coeliac disease, the highest risk is a close family member with disease. Due to its link to coeliac disease, DQ2 has the highest association of any HLA serotype with autoimmune disease, close to 95% of all coeliacs have DQ2, of that 30% have 2 copies of DQ2. Of the DQ2 homozygotes who eat wheat, lifelong risk is between 20 and 40% for coeliac disease.
The relationship of DQ2 and coeliac disease, however, is complex because there are multiple DQ2 isoforms. The DQ α5β2 (DQ2.5) isoform is strongly associated with CD. This isoform is partially encoded by the DQB1*02 genes in HLA-DQ2 positive individuals. DQB1*0201 is genetically linked to DQA1*0501 forming the DQ2.5 haplotype that encodes both α5 and β2 subunits. The DQ2.5 haplotype confers the single highest genetic risk for disease, however comparable risk can also come from very similar alleles on different haplotypes.
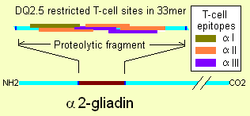
The immunodominant site for DQ2.5 is on α2-gliadin. The site is a protease resistant 33mer that has 6 overlapping DQ2.5 restricted epitopes. This creates very strong binding of T-cells for DQ2.5-33mer complexes. DQ2.5 binds gliadin, but the binding is sensitive to deamidation caused by tissue transglutaminase. In almost all case the highest affinity sites of gluten are derived by deamidation. The HLA DQB1*0202 and its linked DQA1* alleles (the DQ2.2 haplotype) do not produce the α5 subunit. While the DQ2.2 heterodimer cannot effectively present α-2 gliadin, it can present other gliadins. In at least 1% of coeliacs DQ2.2 confers adaptive immunity to gliadin permitting coeliac disease.

DQ2.5 and gluten
As mentioned the DQA1*0501:DQB1*0201 haplotype produces DQ2.5cis which by frequency and efficiency in alpha-gliadin presentation is the major factor in adaptive immunity. The isoform, referred to frequently as the DQ2 heterodimer or DQ2 (DQA1*05:DQB1*02) and more recently DQ2.5 can be differentiated from responses from other DQ isoforms, including other DQ2.[10][11] Specifically, that this DQ2 heterodimer is responsible for presenting the α2-gliadin that most effectively stimulates pathogenic T-cells.
The highest risk for coeliac disease is in Western Ireland and overlaps one of three global nodes of the DQ2.5 haplotype in Western Europe. The DQ2.5 haplotype is linked to DR3 and DR3 is not linked to DQ2.2. Therefore, using either serotyping or genotyping, DQ2.5 can be distinguished from DQ2.2 or DQ2.3. The refined studies of risk and immunology suggest that all DQ2 can mediate coeliac disease, but that DQ2.5 is the primary genetic risk factor. A genome wide survey of markers linked to CD, reveals that highest linkage is for a marker within the DQA1*0501 allele of the DQ2.5 haplotype.[12] The association of DQB1*0201 is almost as high. Greatly elevating risk is the ability of the DQ2.5 haplotype encoded isoforms to increase abundance on the cell surface in DQ2.5 double homozygotes. While most people can form two or four different isoforms of DQ. Double homozygotes (of DQA1 and DQB1) can only form DQ2.5cis. This occurs when a person inherits a DQ2.5cis bearing chromosome from each parent. While the frequency of DQ2.5 haplotype is only 4 times higher than the general population, the number of DQ2.5 homozygotes is 10 to 20 times higher than the general population.[3][13] Multiple copies of the DQ2.5 haplotype do not cause apparent increases of severity, DQ2.5/DQ2 increases risk of life threatening complications and more severe histological findings.[14][15] Of the approximately 90% of coeliacs that bear the DQ2.5 isoform only 4% produce DQ2.5 by pair alleles from different haplotypes, this isoform is called DQ2.5trans and differs slightly, one amino acid, from DQ2.5cis.
DQ2.2 and gluten
DQ2.2 does not produce all the necessary subunits to efficiently present the most pathogenic gluten proteins to the immune system. With the DQ2.2 isoform (DQ α2-β2), polar substitutions (amino acids such as asparagine, glutamine, glycine, serine, and threonine) are not bound well to DQ2.2.[16] The gliadin peptides that bind DQ2.5 are enriched in the amino acid glutamine. Since the β2 provides half the structural information for gluten presentation, other haplotypes might provide the rest. Such haplotypes are known to exist and these haplotypes confer different risk on DQ2.2. DQ2.2 however can present less pathogenic epitopes such as proteolytic peptides of gamma-gliadin. This appears to be the mediator of disease in 1% of coeliacs that are homozygotes for DQ2.2.
The DQ2.2/DQ7.5 phenotype. Also called DQ2.5trans in some publications. DQ7.5 haplotype is the DQA1*0505:DQB1*0301 haplotype. The DQA1*0505 allele is similar to the DQA1*0501 allele of the DQ2.5 haplotype. When DQA1*0505 or DQA1*0501 gene products are processed to the cell surface they become α5. The gene products of DQB1*0202 and DQB1*0201 are almost identical and function similarly. As a result one isoform produced by the phenotype of two haplotypes, DQ2.2/DQ7.5, is HLA DQ α5β2. A small percentage of coeliac disease patients have this haplotype. The other 3 isoforms are α2β2(DQ2.2), α2β7 (DQ7.2), and α5β7 (DQ7.5).
DQ2.2/DQ2.5. Random pairing of heterologous DQ alpha and beta isoforms produces 4 different isoforms at 1:1:1:1 ratios. The fraction of DQ2.5 can be 25%. In the case of this phenotype, HLA DQB1*02 alleles are encoded by both chromosome 6 (maternal and paternal derived). Since DQB1*0201 and *0202 function similarly, only two types of isoforms can be produced and the ratio becomes 1:1. This increases the random number of isoforms from 25% to 50% that can cause disease, and as a result increases risk of celiac disease[17][18] and probably increases risk of severe complications such as refractory celiac disease and lymphoma.[14] These partial homozygotes in the Dutch CD population are approximately 20%, as compared to a randomly expected 3% indicating a sevenfold enrichment.
DQ2.2/DQ8. Among DQ8 positive celiacs without DQ2.5, 1/3 bear DQ2.2 haplotype, about 3 fold higher than random expectation.
DQ2.2/DQ2.2 DQ2.2 homozygotes represent about 1.1% of the celiac population, this is not high relative to controls, but it is very high with the DQ2.5(isoform)-,DQ8-,DQ2+ cohort at 30%. The random expectation is much lower.[4] This fraction of coeliacs is important because they can only produce α2β2 and are useful for determining the role of DQ2.2 in coeliac disease.
Juvenile diabetes association
Juvenile diabetes (T1D) has a high association with DQ2.5 and there appears to be link between GSE and early onset male T1D. Anti-tTG antibodies are found elevated in a one-third of T1D patients[19][20] and there are indicators that Triticeae may be involved but the gluten protein is a type of globulin (Glb1).[21] Recent studies indicate a combination of DQ2.5 and DQ8 (both acid peptide presenters) greatly increase the risk of adult onset Type 1 Diabetes and ambiguous type I/II Diabetes.[22][23] HLA-DR3 plays a prominent role in autoimmune diabetes.[24] However, DQ2 presence with DR3 decreases the age of onset and the severity of the autoimmune disorder.
References
- ↑ derived from IMGT/HLA
- ↑ Klitz W, Maiers M, Spellman S et al. (October 2003). "New HLA haplotype frequency reference standards: high-resolution and large sample typing of HLA DR-DQ haplotypes in a sample of European Americans". Tissue Antigens 62 (4): 296–307. doi:10.1034/j.1399-0039.2003.00103.x. PMID 12974796.
- ↑ 3.0 3.1 van Belzen MJ, Koeleman BP, Crusius JB et al. (May 2004). "Defining the contribution of the HLA region to cis DQ2-positive coeliac disease patients". Genes Immun. 5 (3): 215–20. doi:10.1038/sj.gene.6364061. PMID 15014431.
- ↑ 4.0 4.1 Karell K, Louka AS, Moodie SJ et al. (April 2003). "HLA types in celiac disease patients not carrying the DQA1*05-DQB1*02 (DQ2) heterodimer: results from the European Genetics Cluster on Celiac Disease". Hum. Immunol. 64 (4): 469–77. doi:10.1016/S0198-8859(03)00027-2. PMID 12651074.
- ↑ Taylor C, Smith S, Morgan C, Stephenson S, Key T, Srinivasan M, Cunningham E, Watson P (2000). "HLA and Mooren's ulceration.". Br J Ophthalmol 84 (1): 72–5. doi:10.1136/bjo.84.1.72. PMC 1723219. PMID 10611103.
- ↑ Weinshenker B, Santrach P, Bissonet A, McDonnell S, Schaid D, Moore S, Rodriguez M (1998). "Major histocompatibility complex class II alleles and the course and outcome of MS: a population-based study.". Neurology 51 (3): 742–7. doi:10.1212/wnl.51.3.742. PMID 9748020.
- ↑ Ratanachaiyavong S, Lloyd L, Darke C, McGregor A (1993). "MHC-extended haplotypes in families of patients with Graves' disease.". Hum Immunol 36 (2): 99–111. doi:10.1016/0198-8859(93)90112-E. PMID 8096501.
- ↑ Tjernström F, Hellmer G, Nived O, Truedsson L, Sturfelt G (1999). "Synergetic effect between interleukin-1 receptor antagonist allele (IL1RN*2) and MHC class II (DR17,DQ2) in determining susceptibility to systemic lupus erythematosus.". Lupus 8 (2): 103–8. doi:10.1191/096120399678847560. PMID 10192503.
- ↑ Kim C, Quarsten H, Bergseng E, Khosla C, Sollid L (2004). "Structural basis for HLA-DQ2-mediated presentation of gluten epitopes in celiac disease". Proc Natl Acad Sci USA 101 (12): 4175–9. doi:10.1073/pnas.0306885101. PMC 384714. PMID 15020763.
- ↑ Qiao SW, Bergseng E, Molberg O, Jung G, Fleckenstein B, Sollid LM (July 2005). "Refining the rules of gliadin T cell epitope binding to the disease-associated DQ2 molecule in celiac disease: importance of proline spacing and glutamine deamidation". J. Immunol. 175 (1): 254–61. doi:10.4049/jimmunol.175.1.254. PMID 15972656.
- ↑ Vader W, Stepniak D, Kooy Y et al. (October 2003). "The HLA-DQ2 gene dose effect in celiac disease is directly related to the magnitude and breadth of gluten-specific T cell responses". Proc. Natl. Acad. Sci. U.S.A. 100 (21): 12390–5. doi:10.1073/pnas.2135229100. PMC 218768. PMID 14530392.
- ↑ van Heel DA, Franke L, Hunt KA et al. (2007). "A genome-wide association study for celiac disease identifies risk variants in the region harboring IL2 and IL21". Nat. Genet. 39 (7): 827–9. doi:10.1038/ng2058. PMC 2274985. PMID 17558408.
- ↑ Louka AS, Nilsson S, Olsson M et al. (August 2002). "HLA in coeliac disease families: a novel test of risk modification by the 'other' haplotype when at least one DQA1*05-DQB1*02 haplotype is carried". Tissue antigens 60 (2): 147–54. doi:10.1034/j.1399-0039.2002.600205.x. PMID 12392509.
- ↑ 14.0 14.1 Al-Toma A, Goerres MS, Meijer JW, Peña AS, Crusius JB, Mulder CJ (2006). "Human leukocyte antigen-DQ2 homozygosity and the development of refractory celiac disease and enteropathy-associated T-cell lymphoma". Clin. Gastroenterol. Hepatol. 4 (3): 315–9. doi:10.1016/j.cgh.2005.12.011. PMID 16527694.
- ↑ Jores RD, Frau F, Cucca F et al. (2007). "HLA-DQB1*0201 homozygosis predisposes to severe intestinal damage in celiac disease". Scand. J. Gastroenterol. 42 (1): 48–53. doi:10.1080/00365520600789859. PMID 17190762.
- ↑ Vartdal F, Johansen BH, Friede T et al. (1996). "The peptide binding motif of the disease associated HLA-DQ (alpha 1* 0501, beta 1* 0201) molecule". Eur. J. Immunol. 26 (11): 2764–72. doi:10.1002/eji.1830261132. PMID 8921967.
- ↑ Louka A, Nilsson S, Olsson M, Talseth B, Lie B, Ek J, Gudjónsdóttir A, Ascher H, Sollid L (2002). "HLA in coeliac disease families: a novel test of risk modification by the 'other' haplotype when at least one DQA1*05-DQB1*02 haplotype is carried". Tissue Antigens 60 (2): 147–54. doi:10.1034/j.1399-0039.2002.600205.x. PMID 12392509.
- ↑ Vader W, Stepniak D, Kooy Y, Mearin L, Thompson A, van Rood J, Spaenij L, Koning F (2003). "The HLA-DQ2 gene dose effect in celiac disease is directly related to the magnitude and breadth of gluten-specific T cell responses". Proc Natl Acad Sci USA 100 (21): 12390–5. doi:10.1073/pnas.2135229100. PMC 218768. PMID 14530392.
- ↑ Lampasona V, Bonfanti R, Bazzigaluppi E, Venerando A, Chiumello G, Bosi E, Bonifacio E. (1999). "Antibodies to tissue transglutaminase C in type I diabetes.". Diabetologia. 42 (10): 1195–1198. doi:10.1007/s001250051291. PMID 10525659.
- ↑ Bao F, Yu L, Babu S, Wang T, Hoffenberg EJ, Rewers M, and Eisenbarth GS. (1999). "One third of HLA DQ2 homozygous patients with type 1 diabetes express celiac disease-associated transglutaminase autoantibodies.". J Autoimmun. 13 (1): 143–148. doi:10.1006/jaut.1999.0303. PMID 10441179.
- ↑ MacFarlane AJ, Burghardt KM, Kelly J, Simell T, Simell O, Altosaar I, and Scott FW. (2003). "A type 1 diabetes-related protein from wheat (Triticum aestivum). cDNA clone of a wheat storage globulin, Glb1, linked to islet damage.". J Biol Chem. 278 (1): 54–63. doi:10.1074/jbc.M210636200. PMID 12409286.
- ↑ Horton V, Stratton I, Bottazzo G, Shattock M, Mackay I, Zimmet P, Manley S, Holman R, Turner R (1999). "Genetic heterogeneity of autoimmune diabetes: age of presentation in adults is influenced by HLA DRB1 and DQB1 genotypes (UKPDS 43). UK Prospective Diabetes Study (UKPDS) Group.". Diabetologia 42 (5): 608–16. doi:10.1007/s001250051202. PMID 10333055.
- ↑ Bakhtadze E, Borg H, Stenström G, Fernlund P, Arnqvist H, Ekbom-Schnell A, Bolinder J, Eriksson J, Gudbjörnsdottir S, Nyström L, Groop L, Sundkvist G (2006). "HLA-DQB1 genotypes, islet antibodies and beta cell function in the classification of recent-onset diabetes among young adults in the nationwide Diabetes Incidence Study in Sweden.". Diabetologia 49 (8): 1785–94. doi:10.1007/s00125-006-0293-5. PMID 16783473.
- ↑ Eller E, Vardi P, McFann KK et al. (2007). "Differential effects of DRB1(*)0301 and DQA1(*)0501-DQB1(*)0201 on the activation and progression of islet cell autoimmunity". Genes and Immunity 8 (8): 628–33. doi:10.1038/sj.gene.6364425. PMID 17728790.
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||