GPX4
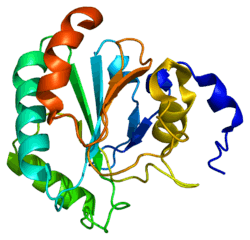
Glutathione peroxidase 4, also known as GPX4, is an enzyme that in humans is encoded by the GPX4 gene.[1] GPX4 is a phospholipid hydroperoxidase that protects cells against membrane lipid peroxidation.
Function
The antioxidant enzyme glutathione peroxidase 4 (GPx4) belongs to the family of glutathione peroxidases, which consists of 8 known mammalian isoenzymes (GPx1-8). Gpx4 catalyzes the reduction of hydrogen peroxide, organic hydroperoxides, and lipid peroxides at the expense of reduced glutathione and functions in the protection of cells against oxidative stress. The oxidized form of glutathione (glutathione disulfide), which is generated during the reduction of hydroperoxides by GPx4, is recycled by glutathione reductase and NADPH/H+. GPx4 differs from the other GPx family members in terms of its monomeric structure, a less restricted dependence on glutathione as reducing substrate, and the ability to reduce lipid-hydroperoxides inside biological membranes.
Mutations in GPX4 cause spondylometaphyseal dysplasia .[2]
Structure
Mammalian GPx1, GPx2, GPx3, and GPx4 (this protein) have been shown to be selenium-containing enzymes, whereas GPx6 is a selenoprotein in humans with cysteine-containing homologues in rodents. In selenoproteins, the 21st amino acid selenocysteine is inserted in the nascent polypeptide chain during the process of translational recoding of the UGA stop codon. GPx4 shares the amino acid motif of selenocysteine, glutamine, and tryptophane (catalytic triad) with other glutathione peroxidases.
Reaction mechanism
GPx4 catalyzes the following reaction:
- 2 glutathione + lipid–hydroperoxide → glutathione disulfide + lipid–alcohol + H2O
This reaction occurs at the selenocysteine within the catalytic center of GPx4. During the catalytic cycle of GPx4, the active selenol (-SeH) is oxidized by peroxides to selenenic acid (-SeOH), which is then reduced with glutathione (GSH) to an intermediate selenodisulfide (-Se-SG). GPx4 is eventually reactivated by a second glutathione molecule, releasing glutathione disulfide (GS-SG).
Subcellular distribution of isoforms
In mouse and rat, three distinct GPx4 isoforms with different subcellular localization are produced through alternative splicing and transcription initiation; cytosolic GPx4, mitochondrial GPx4 (mGPx4), and nuclear GPx4 (nGPx4). Cytosolic GPx4 has been identified as the only GPx4 isoform being essential for embryonic development and cell survival. The GPx4 isoforms mGPx4 and nGPx4 have been implicated in spermatogenesis and male fertility.[3] In humans, experimental evidence for alternative splicing exists; alternative transcription initiation and the cleavage sites of the mitochondrial and nuclear transit peptides need to be experimentally verified.[4]
Animal models
Knockout mice of GPX4 die at embryonic day 8[5][6] and conditional inducible deletion in adult mice (neurons) results in degeneration and death in less than a month.[7] Targeted disruption of the mitochondrial GPx4 isoform (mGPx4) caused infertility in male mice and disruption of the nuclear GPx4 isoform (nGPx4) reduced the structural stability of sperm chromatin, yet both knockout mouse models (for mGPx4 and nGPx4) were fully viable. Surprisingly, knockout of GPX4 heterozygously in mice (GPX4+/−) increases their median life span.[8] Knockout studies with GPx1, GPx2, or GPx3 deficient mice showed that cytosolic GPx4 is so far the only glutathione peroxidase that is indispensable for embryonic development and cell survival. As mechanisms to dispose of both hydrogen peroxide and lipid hydroperoxides are essential to life, this indicates that in contrast to the multiple metabolic pathways that can be utilised to dispose of hydrogen peroxide, pathways for the disposal of lipid hydroperoxides are limited.
While mammals have only one copy of the GPX4 gene, fish have two copies, GPX4a and GPX4b.[9] The GPX4's appear to play a greater role in the fish GPX system than in mammals. For example in fish GPX4 activity contributes to a greater extent to total GPX activity,[10] GPX4a is the most highly expressed selenoprotein mRNA (in contrast to mammals where it is GPX1 mRNA)[11] and GPX4a appears to be highly inducible to changes within the cellular environment, such as changes in methylmercury and selenium status.[12]
References
- ↑ Esworthy RS, Doan K, Doroshow JH, Chu FF (July 1994). "Cloning and sequencing of the cDNA encoding a human testis phospholipid hydroperoxide glutathione peroxidase". Gene 144 (2): 317–8. doi:10.1016/0378-1119(94)90400-6. PMID 8039723.
- ↑ Smith AC, Mears AJ, Bunker R, Ahmed A, MacKenzie M, Schwartzentruber JA, Beaulieu CL, Ferretti E, Majewski J, Bulman DE, Celik FC, Boycott KM, Graham GE (2014). "Mutations in the enzyme glutathione peroxidase 4 cause Sedaghatian-type spondylometaphyseal dysplasia". Journal of Medical Genetics 51 (7): 470–4. doi:10.1136/jmedgenet-2013-102218. PMID 24706940.
- ↑ Schneider M, Förster H, Boersma A, Seiler A, Wehnes H, Sinowatz F, Neumüller C, Deutsch MJ, Walch A, Hrabé de Angelis M, Wurst W, Ursini F, Roveri A, Maleszewski M, Maiorino M, Conrad M (May 2009). "Mitochondrial glutathione peroxidase 4 disruption causes male infertility". FASEB J. 23 (9): 3233–42. doi:10.1096/fj.09-132795. PMID 19417079.
- ↑ "Entrez Gene: GPX4 glutathione peroxidase 4 (phospholipid hydroperoxidase)".
- ↑ Yant LJ, Ran Q, Rao L, Van Remmen H, Shibatani T, Belter JG, Motta L, Richardson A, Prolla TA (February 2003). "The selenoprotein GPX4 is essential for mouse development and protects from radiation and oxidative damage insults". Free Radic. Biol. Med. 34 (4): 496–502. doi:10.1016/S0891-5849(02)01360-6. PMID 12566075.
- ↑ Muller FL, Lustgarten MS, Jang Y, Richardson A, Van Remmen H (2007). "Trends in oxidative aging theories.". Free Radic. Biol. Med. 43 (4): 477–503. doi:10.1016/j.freeradbiomed.2007.03.034. PMID 17640558.
- ↑ Seiler A, Schneider M, Förster H, Roth S, Wirth EK, Culmsee C, Plesnila N, Kremmer E, Rådmark O, Wurst W, Bornkamm GW, Schweizer U, Conrad M (September 2008). "Glutathione peroxidase 4 senses and translates oxidative stress into 12/15-lipoxygenase dependent- and AIF-mediated cell death". Cell Metab. 8 (3): 237–48. doi:10.1016/j.cmet.2008.07.005. PMID 18762024.
- ↑ Ran Q, Liang H, Ikeno Y, Qi W, Prolla TA, Roberts LJ, Wolf N, Van Remmen H, VanRemmen H, Richardson A (2007). "Reduction in glutathione peroxidase 4 increases life span through increased sensitivity to apoptosis". J. Gerontol. A Biol. Sci. Med. Sci. 62 (9): 932–42. doi:10.1093/gerona/62.9.932. PMID 17895430.
- ↑ Mariotti M, Ridge PG, Zhang Y, Lobanov AV, Pringle TH, Guigo R, Hatfield DL, Gladyshev VN (2012). "Composition and evolution of the vertebrate and mammalian selenoproteomes". PLOS one 7 (3): e33066. doi:10.1371/journal.pone.0033066. PMID 22479358.
- ↑ Grim JM, Hyndman KA, Kriska T, Girotti AW, Crockett EL (2011). "Relationship between oxidizable fatty acid content and level of antioxidant glutathione peroxidases in marine fish". The Journal of experimental biology 214 (22): 3751–3759. doi:10.1242/jeb.058214. PMID 22031739.
- ↑ Zheng W, Xu H, Lam SH, Luo H, Karuturi RK, Gong Z (2013). "Transcriptomic analyses of sexual dimorphism of the zebrafish liver and the effect of sex hormones". PLOS one 8 (1): e53562. doi:10.1371/journal.pone.0053562. PMID 23349717.
- ↑ Penglase S, Hamre K, Ellingsen S (2014). "Selenium prevents downregulation of antioxidant selenoprotein genes by methylmercury". Free Radical Biology and Medicine 75: 95–104. doi:10.1016/j.freeradbiomed.2014.07.019. PMID 25064324.
Further reading
- Nakagawa Y (2005). "Role of mitochondrial phospholipid hydroperoxide glutathione peroxidase (PHGPx) as an antiapoptotic factor.". Biol. Pharm. Bull. 27 (7): 956–60. doi:10.1248/bpb.27.956. PMID 15256721.
- Esworthy RS, Doan K, Doroshow JH, Chu FF (1994). "Cloning and sequencing of the cDNA encoding a human testis phospholipid hydroperoxide glutathione peroxidase.". Gene 144 (2): 317–8. doi:10.1016/0378-1119(94)90400-6. PMID 8039723.
- Maruyama K, Sugano S (1994). "Oligo-capping: a simple method to replace the cap structure of eukaryotic mRNAs with oligoribonucleotides.". Gene 138 (1–2): 171–4. doi:10.1016/0378-1119(94)90802-8. PMID 8125298.
- Chu FF (1994). "The human glutathione peroxidase genes GPX2, GPX3, and GPX4 map to chromosomes 14, 5, and 19, respectively". Cytogenet. Cell Genet. 66 (2): 96–8. doi:10.1159/000133675. PMID 8287691.
- Bonaldo MF, Lennon G, Soares MB (1997). "Normalization and subtraction: two approaches to facilitate gene discovery". Genome Res. 6 (9): 791–806. doi:10.1101/gr.6.9.791. PMID 8889548.
- Suzuki Y, Yoshitomo-Nakagawa K, Maruyama K, Suyama A, Sugano S (1997). "Construction and characterization of a full length-enriched and a 5'-end-enriched cDNA library". Gene 200 (1–2): 149–56. doi:10.1016/S0378-1119(97)00411-3. PMID 9373149.
- Opalenik SR, Ding Q, Mallery SR, Thompson JA (1998). "Glutathione depletion associated with the HIV-1 TAT protein mediates the extracellular appearance of acidic fibroblast growth factor". Arch. Biochem. Biophys. 351 (1): 17–26. doi:10.1006/abbi.1997.0566. PMID 9501919.
- Kelner MJ, Montoya MA (1998). "Structural organization of the human selenium-dependent phospholipid hydroperoxide glutathione peroxidase gene (GPX4): chromosomal localization to 19p13.3". Biochem. Biophys. Res. Commun. 249 (1): 53–5. doi:10.1006/bbrc.1998.9086. PMID 9705830.
- Ursini F, Heim S, Kiess M, Maiorino M, Roveri A, Wissing J, Flohé L (1999). "Dual function of the selenoprotein PHGPx during sperm maturation". Science 285 (5432): 1393–6. doi:10.1126/science.285.5432.1393. PMID 10464096.
- Choi J, Liu RM, Kundu RK, Sangiorgi F, Wu W, Maxson R, Forman HJ (2000). "Molecular mechanism of decreased glutathione content in human immunodeficiency virus type 1 Tat-transgenic mice". J. Biol. Chem. 275 (5): 3693–8. doi:10.1074/jbc.275.5.3693. PMID 10652368.
- Richard MJ, Guiraud P, Didier C, Seve M, Flores SC, Favier A (2001). "Human immunodeficiency virus type 1 Tat protein impairs selenoglutathione peroxidase expression and activity by a mechanism independent of cellular selenium uptake: consequences on cellular resistance to UV-A radiation". Arch. Biochem. Biophys. 386 (2): 213–20. doi:10.1006/abbi.2000.2197. PMID 11368344.
- Yagi K, Komura S, Ohishi N (2003). "Expression of human phospholipid hydroperoxide glutathione peroxidase". Methods Mol. Biol. 196: 195–9. doi:10.1385/1-59259-274-0:195. ISBN 1-59259-274-0. PMID 12152199.
|chapter=ignored (help) - Foresta C, Flohé L, Garolla A, Roveri A, Ursini F, Maiorino M (2003). "Male fertility is linked to the selenoprotein phospholipid hydroperoxide glutathione peroxidase". Biol. Reprod. 67 (3): 967–71. doi:10.1095/biolreprod.102.003822. PMID 12193409.
- Borchert A, Savaskan NE, Kuhn H (2003). "Regulation of expression of the phospholipid hydroperoxide/sperm nucleus glutathione peroxidase gene. Tissue-specific expression pattern and identification of functional cis- and trans-regulatory elements". J. Biol. Chem. 278 (4): 2571–80. doi:10.1074/jbc.M209064200. PMID 12427732.
- Villette S, Kyle JA, Brown KM, Pickard K, Milne JS, Nicol F, Arthur JR, Hesketh JE (2003). "A novel single nucleotide polymorphism in the 3' untranslated region of human glutathione peroxidase 4 influences lipoxygenase metabolism". Blood Cells Mol. Dis. 29 (2): 174–8. doi:10.1006/bcmd.2002.0556. PMID 12490284.
- Maiorino M, Bosello V, Ursini F, Foresta C, Garolla A, Scapin M, Sztajer H, Flohe L (2004). "Genetic variations of gpx-4 and male infertility in humans". Biol. Reprod. 68 (4): 1134–41. doi:10.1095/biolreprod.102.007500. PMID 12606444.
- Wang HP, Schafer FQ, Goswami PC, Oberley LW, Buettner GR (2003). "Phospholipid hydroperoxide glutathione peroxidase induces a delay in G1 of the cell cycle". Free Radic. Res. 37 (6): 621–30. doi:10.1080/1071576031000088283. PMID 12868489.
- Chen CJ, Huang HS, Chang WC (2003). "Depletion of phospholipid hydroperoxide glutathione peroxidase up-regulates arachidonate metabolism by 12S-lipoxygenase and cyclooxygenase 1 in human epidermoid carcinoma A431 cells". FASEB J. 17 (12): 1694–6. doi:10.1096/fj.02-0847fje. PMID 12958179.
- Sneddon AA, Wu HC, Farquharson A, Grant I, Arthur JR, Rotondo D, Choe SN, Wahle KW (2004). "Regulation of selenoprotein GPx4 expression and activity in human endothelial cells by fatty acids, cytokines and antioxidants". Atherosclerosis 171 (1): 57–65. doi:10.1016/j.atherosclerosis.2003.08.008. PMID 14642406.
| |||||||||

| |||||||||||||||||