Eodiscina
| Eodiscina Temporal range: Lower and Middle Cambrian | |
|---|---|
 | |
| Eodiscus punctatus, collected from the Plutonites hicksi-zone, Nine Wells Shale, Wales, Middle Cambrian (Menevia), 8mm, head disarticulated from the body | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Arthropoda |
| Class: | Trilobita |
| Order: | Agnostida |
| Suborder: | Eodiscina Kobayashi, 1939 |
| Superfamily: | Eodiscoidea Raymond, 1913 |
| Families | |
| |
Eodiscina is a suborder of trilobites, a well known group of marine arthropods. The Eodiscina first developed near the end of the Lower Cambrian period (late Atdabanian) and went extinct at the end of the Middle Cambrian. They are small or very small, and have a thorax of two or three segments.[1] Eodiscina includes six families classified under one superfamily, Eodiscoidea.
Taxonomy
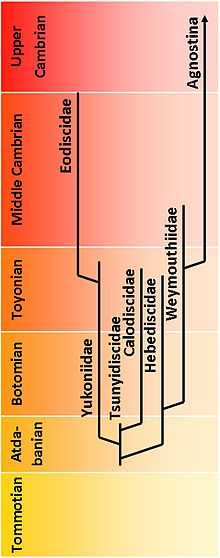
The Eodiscina are mostly considered the more primitive suborder of the Agnostida, and the Agnostina the more advanced.[3] Some scholars do not consider the Agnostina true trilobites, and consequently rejected the idea that they were related to the Eodiscina.[1][4] Consequently, these scientists have proposed to elevate the group to ordinal level, which would thus be called Eodiscida Kobayashi, 1939.[5]
Origin
The oldest known eodiscoid is Tsunyidiscus. The glabella of Tsunyidiscus is extremely similar to that of Dipharus clarki, and distinct from all other eodiscoids. D. clarki is thought to represent an immature stage of the redlichioid Bulaiaspis rather than an eodiscoid. This is because of the dominant palpebroocular ridges, extremely long librigenae, and free pleural tips on the pygidium of variable numbers of segments. In short: Tsunyidiscus has probably developed through paedomorphosis from Bulaiaspis. Alternatively Sinodiscus changyangensis has been considered the most primitive of the Eodiscina.
Many characters that are divergent from other trilobites may be explained by paedomorphosis, such as the small number of thorax segments and proparian facial sutures, that occur in all trilobites during larval development, while the Eodiscoid abatochroal eye is not unlike early development stages of the holochoal eye, where lenses are also separated.[1]
Relations within the Agnostida
Three lineages are thought to have evolved from the monotypical family Tsunyidiscidae. First the Calodiscidae. Second the Yukoniidae, who sprouted the Eodiscidae. And finally the Hebediscidae, that themselves gave rise to the Weymouthiidae, which contain Tannudiscus, the probable ancestor of the peronopsid genus Archaeagnostus,[6] the earliest of the Agnostina.[1]
Differences with some other simplified trilobites
Agnostina and a few other trilobites also have very few thorax segments. All Agnostina lack eyes and only have two thorax segments, characters they share with some of the later Eodiscina. Eodiscina however always have an articulating half-ring. This seals the opening in the axis between cephalon and the anterior thorax segment that is created when the animal was enrolled. Agnostida do not have an articulating half-ring, resulting in an opening between the thorax and the cephalon when enrolled, called cephalothoracic aperture. Thoracocare (Corynexochida) has only two thoracic segments at maturity, but has a very wide subquadrate glabella that touches the frontal border along its entire width and lacks a pygidial border.[4][7] Taklamakania, Pseudampyxina and Nanshanaspis (Raphiophoridae) have 3 thorax segments, but with subtriangular cephalon and pygidium, genal spines, and Taklamakania with a frontal glabellar spine.[8]
Description

Like other Agnostida, the body of Eodiscina is diminutive, the headshield (or cephalon) and tailshield (or pygidium) are of approximately same size, with 2 or 3 thoracic segments in-between, each consisting of a horizontal inner portion that abruptly passes into an inclined outer portion (fulcrate). The cephalon has a deeply parabolic outline and maximum width (tr.) usually anterior to genal angle. The border is convex. The segments of the glabella are commonly fused, and widest at the back-end of the cephalon. The palate (or hypostome), a calcified plate on the underside of the cephalon, is not connected to the frontal border (a state called natant, meaning “floating”). A rostral plate is lacking or uncalcified. With or without eyes. Rupture lines (or sutures) in the headshield, which in most other trilobites assists the periodic moulting (or ecdysis), are present in early Eodiscina, but are lost in some later representatives. When present, the facial sutures are proparian. With an articulating half ring on the anterior thorax segment. The outline of the pygidium is closely matching that of the cephalon. The pygidium is usually segmented, with the axis extending close to the border, and in some species the pleural region is segmented. The calcified first stage larvae (called protaspis) are known in a few species.[1]
Eye
If present, the eyes of Eodiscina are unique among trilobites. They have up to 70 small lenses per eye, not touching but separated from each other by the so-called interlensar sclera. Each lens has an individual cornea, that is limited to the lens surface. The interlensar sclera is not deeper than the lenses. This type of eye is called abatochroal. A second type of eye, called schizochroal, is unique to the Phacopina suborder, and has up to several hundreds of relatively large lenses (0.05-0.5 mm), that do not touch either, but the corneal membrane extends into the sclera outside the lens, and the sclera is deeper than the lens. The most usual type of trilobite eye, called holochroal, has usually thousands of small hexagonal lenses without an interlensar sclera, and a common cornea.[9] Many later Eodiscina however, lost their visual organs altogether, and this may have resulted from living in an environment with very low visibility.[1]
References
- ↑ 1.0 1.1 1.2 1.3 1.4 1.5 Whittington, H. B. et al. Part O, Treatise on Invertebrate Paleontology. Revised, Volume 1 – Trilobita – Introduction, Order Agnostida, Order Redlichiida. 1997
- ↑ Jell, P.A. (1975). "Australian Middle Cambrian Eodiscoids with a review of the superfamily". Palaeontographica Abteilung A 150: 1–97. cited in W.B. Wittington et al., eds. (1993). Treatise on Invertebrate Paleontology, Part O., Revised.
- ↑ Cotton, T.J.; Fortey, R.A. (2005). "5. Comparative morphology and relationships of the Agnostida". In Koenemann, S. & Jenner, R. Crustacean Issues 16, Crustacea and Arthropod Relationships. Boca Raton: CRC Press.
- ↑ 4.0 4.1 Shergold, John H. (1991). "Protaspid and early meraspid growth stages of the eodiscoid trilobite Pagetia ocellata Jell, and their implications for classification". Alcheringa: An Australasian Journal of Palaeontology 15 (1): 65–86. doi:10.1080/03115519108619010.
- ↑ Westrop, S.R.; Landing, E. (2012). "Lower Cambrian (Branchian) eodiscoid trilobites from the lower Brigus formation, Avalon Peninsula, Newfoundland, Canada". Memoirs of the Association of Australasian Palaeontologists 42: 209–262.
- ↑ Naimark, E.B. (2012). "Hundred species of the Genus Peronopsis Hawle et Corda, 1847". Paleontological Journal 46 (9): 945–1057. doi:10.1134/S0031030112090018.
- ↑ S.M. Gon III. "Evolutionary Trends in Trilobites". Retrieved 14 February 2013.
- ↑ Zhou, Z.; Webby, B.D.; Yuan, W. (1995). "Ordovician trilobites from the Yingan Formation of northwestern Tarim, Xinjiang, northwestern China". Alcheringa: An Australasian Journal of Palaeontology 19 (1): 47–72. doi:10.1080/03115519508619098.
- ↑ Sam Gon III. "The Trilobite Eye". Retrieved 16 November 2012.