Elaphrosaurus
| Elaphrosaurus Temporal range: Late Jurassic, 154–150Ma | |
|---|---|
 | |
| Holotype skeleton mounted (skull, hands and other elements speculative), Museum für Naturkunde Berlin | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Dinosauria |
| Suborder: | Theropoda |
| Clade: | Averostra |
| Clade: | †Ceratosauria |
| Genus: | †Elaphrosaurus Janensch, 1920 |
| Species: | † E. bambergi |
| Binomial name | |
| Elaphrosaurus bambergi Janensch, 1920 | |
Elaphrosaurus (pron.:ell-AH-fro-SAWR-us) is a genus of ceratosaurian theropod dinosaur that lived approximately 154 to 150 million years ago during the later part of the Jurassic Period in what is now Tanzania in Africa. Elaphrosaurus was a medium-sized, but lightly built, bipedal, carnivore, that could grow up to 6.2 m (20 ft) long. Morphologically, this dinosaur is significant in two ways. First, it has a relatively long trunk but is very shallow-chested for a theropod of its size. Second, it has very short hindlimbs when compared to its relatively long trunk. Phylogenetic analysis indicates that this genus is likely a ceratosaur, and earlier suggestions that it is a late surviving coelophysoid have been examined but generally dismissed.
Etymology
The genus name Elaphrosaurus is derived from the Greek words elaphro (ελαφρός) meaning "light" as in "light-weight", a reference to its slender frame and "sauros" (σαυρος) meaning "lizard";[1] thus, "lightweight lizard". Elaphrosaurus was described and named by Werner Janensch in 1920 and the type species is Elaphrosaurus bambergi.
Description

Elaphrosaurus was long and slender, with a long neck. What is known about Elaphrosaurus mostly comes from a single nearly complete skeleton and no skull has been found. It was distinctive among theropods for being short-legged for its length. Paul (1988) noted that this was the longest-trunked and shallowest-chested theropod that he has examined.[2] Elaphrosaurus was about 6.2 m (20 ft) long, 1.46 m (5 ft) tall at the hip, and weighed about 210 kilograms (463 pounds) .[2] The tibia (shin bone) of Elaphrosaurus, measured 608 mm was considerably longer than its femur (thigh bone) that measured 520 mm, which indicates that it could probably run very fast.[3] Its long tail ended with a rare downward bend which may be unrelated to taphonomy.
Classification
Elaphrosaurus was first described as a coelurid.[4] At the time, Coeluridae was a wastebasket taxon for small theropods. Then, Elaphrosaurus was placed in the family Ornithomimidae by Nopcsa (1928c) because of its light frame and the fact that its humerus is straight and slender.[5][6][7] Upon closer examination its limbs approximate those of Coelophysis.[2] Barsbold, Maryanska and Osmolska (1990) and other researchers classified it as an ornithomimid[8] More recent work by Carrano and Sampson (2008) and Carrano et al. (2012) assign this genus to Ceratosauria.[9][10] It is now believed that Limusaurus is its closest relative.[11]
The following is a cladogram based on the phylogenetic analysis conducted by Diego Pol and Oliver W. M. Rauhut in 2012,[11] showing the relationships of Elaphrosaurus:
| Ceratosauria |
| |||||||||||||||||||||||||||||||||||||||||||||
| |
Nomina dubia (dubious names)
The following material was assigned to Elaphrosaurus over the years, but further study revealed that these assignments were dubious:
- Elaphrosaurus iguidiensis, was described by Lapparent in 1960,[12] and the material was collected in Algeria, Libya and Niger in Early Cretaceous sediments. The material consists of over 40 teeth, a manual ungual, eight caudal vertebrae, a distal femur fragment, and a complete tibia measuring 350 mm. These specimens originated in three different localities and do not appear to belong to the same species.
- Elaphrosaurus gautieri, was first described by Lapparent in 1960,[12] and the material was collected at the Tiouraren Formation in Niger in Middle-Late Jurassic sediments. This material, a complete neck vertebra, has since been renamed Spinostropheus gautieri by Sereno et al. (2004).[13]
- Elaphrosaurus philtippettensis, was erected by Pickering[14] in 1995 based on USNM 5737, which consists of a tibia, a humerus, some metatarsals, and the distal portion of a fragmentary pubic bones recovered from the Morrison Formation of Colorado. Further research by Carpenter et al. (2005) suggested that the fossils are not ceratosaurian and are likely referable to the coelurid theropod Tanycolagreus.
- Elaphrosaurus agilis, was described Dale Russel in 1972, based on a pair of fused pubic bones that O.C. Marsh had earlier named Coelurus agilis, believing that this specimen represented a much larger version of the type specimen, Coelurus fragilis. John Ostrom (1980) confirmed Charles Gilmore's earlier position that Coelurus agilis was synonymous with Coelurus fragilis. This means that Elaphrosaurus agilis is actually the same animal as Coelurus fragilis.
- Elaphrosaurus sp. USNM 8415, was discovered in 1883 and first referred to the ornithopod Dryosaurus. It was later referred to Elaphrosaurus by Galton in 1982, based on remains recovered at the Morrison Formation of Colorado. This material, which is clearly ceratosaurian, does not bare any morphology that specifically ties it to Elaphrosaurus. Current knowledge limits the placement of this material to Ceratosauria incertae sedis.[15]
- Elaphrosaurus sp. DMNH 36284, was referred to this genus by Chure[16] in 2001, based on the proximal portion of a fragmentary right tibia from the Brushy Basin Member of the Morrison Formation. Phylogenetic analysis by Carrano and Sampson (2008) showed that it was not ceratosaurian, but instead resembled the leg bone of an abelisauroid theropod that has yet to be formally described.[15]
Distinguishing anatomical features
A diagnosis is a statement of the anatomical features of an organism (or group) that collectively distinguish it from all other organisms. Some, but not all, of the features in a diagnosis are also autapomorphies. An autapomorphy is a distinctive anatomical feature that is unique to a given organism.
According to Rauhut (2000), Elaphrosaurus can be distinguished based on the following characteristics:
- the cervical vertebrae possess thin latero-ventral laminae, bordering the posterior pleurocoel ventrally
- the cervical vertebrae are strongly concave ventrally, with the ventral margin arching above the mid-height of the anterior articular facet at its highest point
- the brevis fossa of the ilium is extremely widened, so that the brevis shelf forms an almost horizontal lateral flange
- the distal end of the ischium is strongly expanded into a triangular boot
Paleoecology
Provenance and occurrence
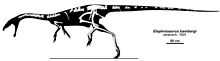
The type specimen of Elaphrosaurus bambergi HMN Gr.S. 38-44 was recovered in the Middle Dinosaur Member of the Tendaguru Formation in Tanzania. The specimen was collected by Werner Janensch, I. Salim, H. Reck, and Parkinson in 1910 in gray, green, red, sandy marl that was deposited during the Kimmeridgian stage of the Jurassic period, approximately 157 to 152 million years ago. This specimen is housed in the collection of the Humboldt Museum in Berlin, Germany.
A related animal, perhaps the same genus, was found in stratigraphic zones 2–4 of the Morrison Formation.[2][17] Few theropod skeletons have been found, most discoveries being fragments.
Fauna and habitat
Studies suggest that the paleoenvironment of the Tendaguru Formation was a marginal marine environment with both non-marine faunal and floral content. The Middle Dinosaur Member of the Tendaguru Formation has yielded the sauropods Giraffatitan, Australodocus, Janenschia, Tornieria and Dicraeosaurus, theropods similar to Allosaurus and Ceratosaurus, the carcharodontosaurid Veterupristisaurus, the stegosaurid Kentrosaurus and the iguanodontian Dysalotosaurus. Dinosaurs shared this paleoenvironment with pterosaurs like Pterodactylus and Rhamphorhynchus, as well as with early mammals. Paul (1988) noted that Elaphrosaurus bambergi was too small to prey on the sauropods and stegosaurs present in its paleoenvironment, and instead, it likely hunted the small and swift ornithopod herbivores.[2]
Ichnology
Dinosaur footprints from the Niger Republic and from Jerusalem were attributed to Elaphrosaurus.[18][19] This assignment is considered inconclusive.
References
- ↑ Liddell, Henry George and Robert Scott (1980). A Greek-English Lexicon (Abridged Edition). United Kingdom: Oxford University Press. ISBN 0-19-910207-4.
- ↑ 2.0 2.1 2.2 2.3 2.4 Paul, Gregory S. (1988). "Genus Elaphrosaurus". Predatory Dinosaurs of the World. New York: Simon & Schuster. pp. 265–266. ISBN 0-671-61946-2.
- ↑ Foster, John (2007). Jurassic West: The Dinosaurs of the Morrison Formation and Their World. Indiana University Press. p. 182. ISBN 978-0-253-34870-8.
- ↑ Janensch, Werner (1920). "Über Elaphrosaurus bambergi und die Megalosaurier aus den Tendaguru–Schichten Deutsch–Ostafrikas". Sitzungsberichte der Gessellschaft Naturforschender Freunde zu Berlin (in German) 1920: 225–235.
- ↑ Galton, 1982. Elaphrosaurus, an ornithomimid dinosaur from the Upper Jurassic of North America and Africa. Paläontologische Zeitschrift. 56, 265−275.
- ↑ Russell, Dale A. (1972). "Ostrich dinosaurs from the Late Cretaceous of western Canada". Canadian Journal of Earth Sciences 9: 375–402. Bibcode:1972CaJES...9..375R. doi:10.1139/e72-031.
- ↑ Nopcsa, F. (1928). The genera of reptiles: Paleobiologica, 1, pp. 163−188.
- ↑ Barsbold, R; Maryanska, T; & Osmolska, H: Oviraptorosauria. Weishampel, D B, Dodson, P, & Osmolska, H, editors: The Dinosauria. University of California Press, Berkeley; 1990.
- ↑ M. T. Carrano and S. D. Sampson. 2008. The phylogeny of Ceratosauria (Dinosauria: Theropoda). Journal of Systematic Palaeontology 6(2):183−236
- ↑ M. T. Carrano, R. B. J. Benson, and S. D. Sampson. 2012. The phylogeny of Tetanurae (Dinosauria: Theropoda). Journal of Systematic Palaeontology 10(2):211–300
- ↑ 11.0 11.1 Diego Pol & Oliver W. M. Rauhut (2012). "A Middle Jurassic abelisaurid from Patagonia and the early diversification of theropod dinosaurs". Proceedings of the Royal Society B: Biological Sciences 279 (1804): 3170–5. doi:10.1098/rspb.2012.0660. PMC 3385738. PMID 22628475.
- ↑ 12.0 12.1 Lapparent, 1960. Les dinosauriens du "Continental intercalaire" du Sahara central. Mem. Soc. Geol. France. 88A 1-57.
- ↑ Sereno, Wilson and Conrad, 2004. New dinosaurs link southern landmasses in the Mid-Cretaceous. Proceedings: Biological Sciences. 71(1546), 1325–1330.
- ↑ Pickering, 1995a. Jurassic Park: Unauthorized Jewish Fractals in Philopatry. A Fractal Scaling in Dinosaurology Project, 2nd revised printing. Capitola, California. 478 pp.
- ↑ 15.0 15.1 Carrano and Sampson, 2008. The phylogeny of Ceratosauria (Dinosauria: Theropoda). Journal of Systematic Palaeontology. 6, 183–236.
- ↑ Chure, 2001. The second record of the African theropod Elaphrosaurus (Dinosauria, Ceratosauria) from the Western Hemisphere. Neues Jahrbuch für Geologie und Paläontologie Monatshefte. 2001(9), 565–576.
- ↑ Foster, J. (2007). "Appendix." Jurassic West: The Dinosaurs of the Morrison Formation and Their World. Indiana University Press. pp. 327–329.
- ↑ M. Avnimelech. 1962. Dinosaur tracks in the Lower Cenomanian of Jerusalem. Nature 196(4851):264
- ↑ Ginsburg, L., Lapparent, A.F. deLoiret, B.and Taquet, P. (1966) Empreintes de pas de Vertebres tetrapodes dans les series continentales a l'Ouest d'Agades (Republique du Niger). Compte Rendu de l'Academie des Sciences, Paris, 263: 28–31.
- Khalaf-von Jaffa, Norman Ali Bassam Ali Taher (2006). "Ornithomimid Dinosaur Tracks from Beit Zeit, West of Jerusalem, Palestine." Gazelle: The Palestinian Biological Bulletin, Number 56, August 2006. pp. 1–7.
- Channell, Michael Benton ; illustrated by Jim; Maddison, Kevin (1988). Dinosaurs : an a-z guide. New York: Derrydale Books. ISBN 978-0517668771.
- Werner Janensch (1925) "Die Coelurosaurier und Theropoden der Tendaguru-Schichten Deutsch-Ostafrikas". (The coelurosaurs and theropods of the Tendaguru Formation, German East Africa). Full Text here: Janensch1925