Default mode network

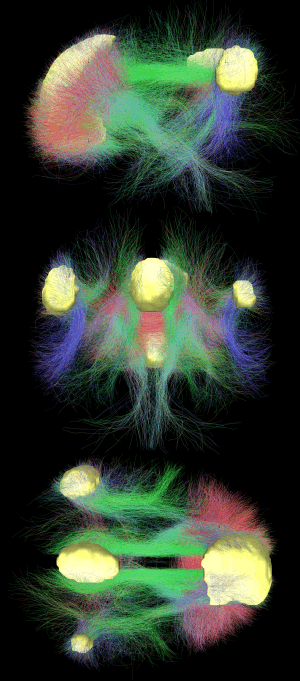
In neuroscience, the default mode network (DMN) is a network of brain regions that are active when the individual is not focused on the outside world and the brain is at wakeful rest. Also called the default network, default state network, or task-negative network, the DMN is characterized by coherent neuronal oscillations at a rate lower than 0.1 Hz (one every ten seconds). During goal-oriented activity, the DMN is deactivated and another network, the task-positive network (TPN) is activated. The DMN may correspond to task-independent introspection, or self-referential thought, while the TPN corresponds to action, and thus perhaps the DMN and TPN may be "considered elements of a single default network with anti-correlated components".[4]
Anatomy
The default mode network is an interconnected and anatomically defined[5] brain system that preferentially activates when individuals engage in internal tasks such as daydreaming, envisioning the future, retrieving memories, and gauging others' perspectives.[5] It is negatively correlated with brain systems that focus on external visual signals. Its subsystems include part of the medial temporal lobe for memory, part of the medial prefrontal cortex for theory of mind, and the posterior cingulate cortex for integration,[5] along with the adjacent ventral precuneus[6] and the medial, lateral and inferior parietal cortex. It has been shown that the default mode network exhibits the highest overlap in its structural and functional connectivity, which suggests that the structural architecture of the brain may be built in such a way that this particular network is activated (by default) as long as no other mental activities are in process (i.e. in a task-absent state).[7] In the infant brain, there is limited evidence of the default network, but default network connectivity is more consistent in children aged 9–12 years, suggesting that the default network undergoes developmental change.[4]
Function
The function of the default mode network remains unclear; however, activity in the network negatively correlates with activity in regions involved in attention and executive function.[8] In humans, the default mode network has been hypothesized to generate spontaneous thoughts during mind-wandering and may relate to creativity.[5][9] Alternatively, default mode activity may represent underlying physiological processes going on in the brain that are unrelated to any particular thought or thoughts.[10] The posterior cingulate gyrus discerns emotional and self-relevant information; this interacts with both the anterior cingulate gyrus, which integrates emotional information with cognition, and the medial prefrontal cortex, which allows for self-reflection and the regulation of emotion and arousal. Attribution of arousal and emotion regulation to the DMN appears to correlate with the experience of people who have experienced long-term trauma and describe feeling 'dead inside' or have a fragmented sense of self or enter dissociative states.
Pathophysiology
The default mode network has been hypothesized to be relevant to disorders including Alzheimer's disease, autism, schizophrenia, depression, chronic pain and others.[5] In particular, a failure to deactivate default network activity during cognitively demanding tasks is associated with autism,[11] overactivity with schizophrenia,[12] and the default network is preferentially attacked by the buildup of beta-amyloid in Alzheimer's disease.[13] Lower connectivity was found across the default network in people who have experienced long term trauma, such as childhood abuse. Among people experiencing posttraumatic stress disorder, lower activation was found in the posterior cingulate gyrus compared to controls.[14] Hyperconnectivity of the default network has been linked to rumination in depression[15] and chronic pain.[16]
Modulation
The default mode network (DMN) may be modulated by the following interventions and processes:
- Acupuncture - Deactivation of the limbic brain areas and the DMN.[17] Tactile control stimulation results in deactivation patterns that are considerably weaker and not statistically significant.[18]
- Meditation - Structural changes in areas of the DMN such as the temporoparietal junction, posterior cingulate cortex, and precuneus have been found in meditation practitioners.[19] There is reduced activation and reduced functional connectivity of the DMN in long-term practitioners.[19]
- Sleeping and resting wakefulness
- Resting wakefulness - Functional connectivity between nodes of the DMN is strong.[20]
- Onset of sleep - Decrease in connectivity between the DMN and the task-positive network.[20]
- Stage N2 of NREM sleep - Decrease in connectivity between the posterior cingulate cortex and medial prefrontal cortex.[20]
- Stage N3 of NREM sleep - Further decrease in connectivity between the PCC and MPFC.[20]
- REM sleep - Possible increase in connectivity between nodes of the DMN.[20]
- Sleep deprivation - Functional connectivity between nodes of the DMN in their resting-state is usually strong, but sleep deprivation results in a decrease in connectivity within the DMN.[21] Recent studies suggest a decrease in connectivity between the DMN and the task-positive network as a result of sleep loss.[22]
- Psychedelic drugs - Reduced blood flow to the PCC and mPFC were observed under the administration of Psilocybin. These two areas are believed to be involved in the DMN.[23]
History
Hans Berger, the inventor of the electroencephalogram, was the first to propose the idea that the brain is constantly busy. In a series of papers published in 1929 he showed that the electrical oscillations detected by his device do not cease even when the subject is at rest. However, his ideas were not taken seriously, and a general perception formed among neurologists that only when a focused activity is performed does the brain (or a part of the brain) become active.[24] In the 1970s, intrinsic oscillatory behavior in vertebrate neurons was observed in cerebellar Purkinje cells, inferior olivary nucleus and thalamus.[25]
Later, experiments by neurologist Marcus E. Raichle's lab at Washington University School of Medicine and other groups showed that the brain's energy consumption is increased by less than 5% of its baseline energy consumption while performing a focused mental task. These experiments showed that the brain is constantly active with a high level of activity even when the person is not engaged in focused mental work. Research thereafter focused on finding the regions responsible for this constant background activity level.[24]
Raichle coined the term "default mode" in 2001 to describe resting state brain function;[26] the concept rapidly became a central theme in neuroscience.[10] In later work, the default mode network was demonstrated by measuring correlated functional activity using resting state fMRI. The brain has other Resting State Networks (RSNs), such as visual and auditory networks.[4]
Tom Eichele, a neuroscientist at the University of Bergen in Norway, as part of an international team of researchers, identified a change in brain scans up to thirty seconds before a mistake, associated with brain activity changes. The researchers suspect the abnormal behavior is the result of the brain attempting to save effort on a task by entering a more restful state.[27][28]
Criticism
The idea of a "default network" is not universally accepted.[29] In 2007 the concept of the default mode was criticized as not being useful for understanding brain function, on the grounds that a simpler hypothesis is that a resting brain actually does more processing than a brain doing certain "demanding" tasks, and that there is no special significance to the intrinsic activity of the resting brain.[30]
References
- ↑ Fair, Damien A.; Cohen, Alexander L.; Power, Jonathan D.; Dosenbach, Nico U. F.; Church, Jessica A.; Miezin, Francis M.; Schlaggar, Bradley L.; Petersen, Steven E. (2009). Sporns, Olaf, ed. "Functional Brain Networks Develop from a 'Local to Distributed' Organization". PLoS Computational Biology 5 (5): e1000381. doi:10.1371/journal.pcbi.1000381. PMC 2671306. PMID 19412534.
- ↑ Horn, Andreas; Ostwald, Dirk; Reisert, Marco; Blankenburg, Felix (2013). "The structural-functional connectome and the default mode network of the human brain". NeuroImage 102: 142–151. doi:10.1016/j.neuroimage.2013.09.069. PMID 24099851.
- ↑ Garrity, A.; Pearlson, G. D.; McKiernan, K.; Lloyd, D.; Kiehl, K. A.; Calhoun, V. D. (2007). "berrant default mode functional connectivity in schizophrenia". Am.J.Psychiatry 164: 450–457. doi:10.1176/appi.ajp.164.3.450.
- ↑ 4.0 4.1 4.2 Broyd, Samantha J.; Demanuele, Charmaine; Debener, Stefan; Helps, Suzannah K.; James, Christopher J.; Sonuga-Barke, Edmund J.S. (2009). "Default-mode brain dysfunction in mental disorders: A systematic review". Neuroscience & Biobehavioral Reviews 33 (3): 279–96. doi:10.1016/j.neubiorev.2008.09.002. PMID 18824195.
- ↑ 5.0 5.1 5.2 5.3 5.4 Buckner, R. L.; Andrews-Hanna, J. R.; Schacter, D. L. (2008). "The Brain's Default Network: Anatomy, Function, and Relevance to Disease". Annals of the New York Academy of Sciences 1124 (1): 1–38. doi:10.1196/annals.1440.011. PMID 18400922.
- ↑ Zhang S, Li CS; Li (February 2012). "Functional connectivity mapping of the human precuneus by resting state fMRI". NeuroImage 59 (4): 3548–3562. doi:10.1016/j.neuroimage.2011.11.023. PMC 3288461. PMID 22116037.
- ↑ Horn A, Ostwald D, Reisert M, Blankenburg F; Blankenburg (2013). "The structural-functional connectome and the default mode network of the human brain". NeuroImage 13: 1053–8119. doi:10.1016/j.neuroimage.2013.09.069. PMID 24099851.
- ↑ Fornito, A; Harrison, B. J.; Zalesky, A; Simons, J. S. (2012). "Competitive and cooperative dynamics of large-scale brain functional networks supporting recollection". Proceedings of the National Academy of Sciences 109 (31): 12788–93. doi:10.1073/pnas.1204185109. PMC 3412011. PMID 22807481.
- ↑ Baird, B.; Smallwood, J.; Mrazek, M. D.; Kam, J. W. Y.; Franklin, M. S.; Schooler, J. W. (2012). "Inspired by Distraction: Mind Wandering Facilitates Creative Incubation". Psychological Science 23 (10): 1117–1122. doi:10.1177/0956797612446024. PMID 22941876.
- ↑ 10.0 10.1 Raichle, Marcus E.; Snyder, Abraham Z. (2007). "A default mode of brain function: A brief history of an evolving idea". NeuroImage 37 (4): 1083–90. doi:10.1016/j.neuroimage.2007.02.041. PMID 17719799.
- ↑ Kennedy, DP; Redcay, E; Courchesne, E (2006). "Failing to deactivate: resting functional abnormalities in autism". Proc. Natl. Acad. Sci. USA 103 (21): 8275–8280. doi:10.1073/pnas.0600674103. PMC 1472462. PMID 16702548.
- ↑ Whitfield-Gabrieli, S.; Thermenos, H. W.; Milanovic, S.; Tsuang, M. T.; Faraone, S. V.; McCarley, R. W.; Shenton, M. E.; Green, A. I. et al. (2009). "Hyperactivity and hyperconnectivity of the default network in schizophrenia and in first-degree relatives of persons with schizophrenia". Proceedings of the National Academy of Sciences 106 (4): 1279–84. doi:10.1073/pnas.0809141106. PMC 2633557. PMID 19164577.
- ↑ Kolata G (2010-12-13). "Insights give hope for new attack on Alzheimer's". New York Times. Retrieved 2010-12-14.
- ↑ Dr. Ruth Lanius, Brain Mapping conference, London, November 2010)
- ↑ Zhu, X; Wang, X; Xiao, J; Liao, J; Zhong, M; Wang, W; Yao, S (2012). "Evidence of a dissociation pattern in resting-state default mode network connectivity in first-episode, treatment-naive major depression patients". Biological Psychiatry 71 (7): 611–7. doi:10.1016/j.biopsych.2011.10.035. PMID 22177602.
- ↑ Kucyi, A; Moayedi, M; Weissman-Fogel, I; Goldberg, M. B.; Freeman, B. V.; Tenenbaum, H. C.; Davis, K. D. (2014). "Enhanced medial prefrontal-default mode network functional connectivity in chronic pain and its association with pain rumination". Journal of Neuroscience 34 (11): 3969–75. doi:10.1523/JNEUROSCI.5055-13.2014. PMID 24623774.
- ↑ Huang, Wenjing; Pach, Daniel; Napadow, Vitaly; Park, Kyungmo; Long, Xiangyu; Neumann, Jane; Maeda, Yumi; Nierhaus, Till; Liang, Fanrong; Witt, Claudia M.; Harrison, Ben J. (9 April 2012). "Characterizing Acupuncture Stimuli Using Brain Imaging with fMRI - A Systematic Review and Meta-Analysis of the Literature". PLOS ONE 7 (4): e32960. doi:10.1371/journal.pone.0032960.
- ↑ Chae, Younbyoung; Chang, Dong-Seon; Lee, Soon-Ho; Jung, Won-Mo; Lee, In-Seon; Jackson, Stephen; Kong, Jian; Lee, Hyangsook; Park, Hi-Joon; Lee, Hyejung; Wallraven, Christian. "Inserting Needles Into the Body: A Meta-Analysis of Brain Activity Associated With Acupuncture Needle Stimulation". The Journal of Pain 14 (3): 215–222. doi:10.1016/j.jpain.2012.11.011.
- ↑ 19.0 19.1 Fox, Kieran C.R.; Nijeboer, Savannah; Dixon, Matthew L.; Floman, James L.; Ellamil, Melissa; Rumak, Samuel P.; Sedlmeier, Peter; Christoff, Kalina. "Is meditation associated with altered brain structure? A systematic review and meta-analysis of morphometric neuroimaging in meditation practitioners". Neuroscience & Biobehavioral Reviews 43: 48–73. doi:10.1016/j.neubiorev.2014.03.016.
- ↑ 20.0 20.1 20.2 20.3 20.4 Picchioni, Dante; Duyn, Jeff H.; Horovitz, Silvina G. "Sleep and the functional connectome". NeuroImage (journal) 80: 387–396. doi:10.1016/j.neuroimage.2013.05.067.
- ↑ McKenna, Benjamin S.; Eyler, Lisa T. "Overlapping prefrontal systems involved in cognitive and emotional processing in euthymic bipolar disorder and following sleep deprivation: A review of functional neuroimaging studies". Clinical Psychology Review 32 (7): 650–663. doi:10.1016/j.cpr.2012.07.003. PMC 3922056. PMID 22926687.
- ↑ Basner, Mathias; Rao, Hengyi; Goel, Namni; Dinges, David F. "Sleep deprivation and neurobehavioral dynamics". Current Opinion in Neurobiology 23 (5): 854–863. doi:10.1016/j.conb.2013.02.008.
- ↑ Carhart-Harris, Robin L.; Erritzoe, David; Williams, Tim; Stone, James M.; Reed, Laurence J.; Colasanti, Alessandro; Tyacke, Robin J.; Leech, Robert; Malizia, Andrea L.; Murphy, Kevin; Evans, John; Feilding, Amanda; Wise, Richard G.; Nutt, David J. "Neural correlates of the psychedelic state as determined by fMRI studies with psilocybin". PNAS 109 (6): 2138–2143. doi:10.1073/pnas.1119598109. PMC 3277566. PMID 22308440.
- ↑ 24.0 24.1 Raichle, Marcus (March 2010). "The Brain's Dark Energy". Scientific American: 44–49.
- ↑ Llinas, R. R. (2014). "Intrinsic electrical properties of mammalian neurons and CNS function: a historical perspective". Front Cell Neurosci 8: 320. doi:10.3389/fncel.2014.00320. PMC 4219458. PMID 25408634.
- ↑ Raichle, M. E.; MacLeod, AM; Snyder, AZ; Powers, WJ; Gusnard, DA; Shulman, GL (2001). "Inaugural Article: A default mode of brain function". Proceedings of the National Academy of Sciences 98 (2): 676–82. doi:10.1073/pnas.98.2.676. PMC 14647. PMID 11209064.
- ↑ Eichele, Tom; Debener, Stefan; Calhoun, Vince D.; Specht, Karsten; Engel, Andreas K.; Hugdahl, Kenneth; Cramon, D. Yves von; Ullsperger, Markus (April 21, 2008). "Prediction of human errors by maladaptive changes in event-related brain networks". Proceedings of the National Academy of Sciences 105 (16): 6173–8. doi:10.1073/pnas.0708965105. PMC 2329680. PMID 18427123.
- ↑ Choi, Charles Q. (April 21, 2008). "Mind-Reading Hat Could Prevent Brain Farts". LiveScience. Retrieved May 18, 2014.
- ↑ Fair, D. A.; Cohen, A. L.; Dosenbach, N. U. F.; Church, J. A.; Miezin, F. M.; Barch, D. M.; Raichle, M. E.; Petersen, S. E.; Schlaggar, B. L. (2008). "The maturing architecture of the brain's default network". Proceedings of the National Academy of Sciences 105 (10): 4028–32. doi:10.1073/pnas.0800376105. PMC 2268790. PMID 18322013.
- ↑ Morcom, Alexa M.; Fletcher, Paul C. (2007). "Does the brain have a baseline? Why we should be resisting a rest". NeuroImage 37 (4): 1073–1082. doi:10.1016/j.neuroimage.2006.09.013.
External links
- Healy, Melissa "An idle brain may be the self's workshop", Los Angeles Times, August 30, 2010
- Healy, Melissa "Brain's default mode network may hold key to better psychiatric diagnoses", Los Angeles Times, August 30, 2010