Basal (phylogenetics)
In phylogenetics, basal is the direction of the base (or root) of a rooted phylogenetic tree or cladogram. Clade C may be described as basal within a larger clade D if its root is directly linked (adjacent) to the root of D. If C is the only clade of a given taxonomic rank that is basal within D, C may be described as the basal taxon of that rank within D. While there must always be two or more equally basal clades sprouting from the root of every cladogram, those clades may differ widely in rank[n 1] and/or species diversity. Greater diversification may be associated with more evolutionary innovation, but ancestral characters should not be imputed to the members of a less species-rich basal clade without additional evidence, as there can be no assurance such an assumption is valid.
In general, clade A is more basal than clade B if B is a subgroup of the sister group of A. Within large groups, "basal" may be used more loosely to mean 'closer to the root than the great majority of', and in this context terminology such as "very basal" may arise.
Examples
A basal group forms a sister group to the rest of the larger clade, such as in the following example:
| |
| ||||||||||||||||||
| |
The term basal can be correctly applied to clades of organisms, but not to lineages or to individual traits possessed by the organisms—although it may be misused in these ways in technical literature. While "basal" applies to clades, characters or traits are usually considered derived if they are absent in a basal group, but present in other groups. This assumption only holds true if the basal group is a good analogy for the last common ancestor of the group. "Ancestral" or "basal" are preferred to "primitive", which may carry false connotations of inferiority or a lack of complexity.
The flowering plant family Amborellaceae, restricted to New Caledonia in the southwestern Pacific,[n 2] is a basal clade of extant angiosperms, containing the most basal species, genus, family and order within the group (out of a total of about 250,000 angiosperm species). While the traits of Amborella trichopoda are regarded as providing significant insight into the evolution of flowering plants, they are a mix of archaic and derived features that have only been sorted out via comparison with other angiosperms and their positions within the phylogenetic tree (the fossil record could potentially also be helpful in this respect, but is absent in this case).[1]
| |
| |||||||||||||||
| |
Within the primate family Hominidae, gorillas (eastern and western) are a sister group to common chimpanzees, bonobos and humans. These five species form a clade, the subfamily Homininae, of which Gorilla is the basal genus.
| |
| ||||||||||||||||||||||||
| |
However, orangutans are a sister group to Homininae and are the basal genus in the family as a whole.
| |
| |||||||||||||||||||||
| |
Subfamilies Homininae and Ponginae are both basal within Hominidae, but given that there are no nonbasal subfamilies in the cladogram it is unlikely the term would be applied to either.
Criticism
Despite the ubiquity of the usage of the term, some systematists believe its application to extant groups is unnecessary and misleading.[2] This is because a pair of sister branches of a phylogenetic tree are equally ranked (in terms of the highest taxonomic ranks of the respective clades) and can be freely rotated about their last common node. The term tends to be applied when one branch (the one deemed "basal") is less diverse than another branch (this being the situation in which one would expect to find basal taxa of lower minimum rank). This raises an equivocal usage for this term in that it also refers to the direction of the root of the tree. As the root represents a hypothetical ancestor, this consequently may carry misleading connotations that the sister group of a more species-rich clade is primitive.[3]
Relevance to biogeographic history
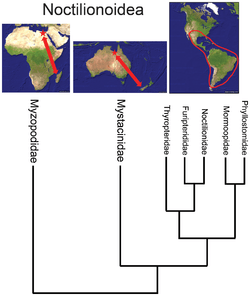
Given that the deepest phylogenetic split in a widely dispersed group is likely to have occurred in its region of origin, identification of the most basal clade(s) in such a group can provide valuable insight into its biogeographic history. For example, extant australidelphian marsupials constitute about 238 species in Australasia and one species (the monito del monte) in South America. The fact that the monito del monte occupies a basal position (the most basal species, genus, family and order) in the superorder is an important clue that its origin was in South America.[4][n 3] Also, while the bat superfamily Noctilionoidea has over 200 species in the Neotropics, two in New Zealand, and two in Madagascar, the basal position of the Malagasy genus and family[5] suggests (in combination with the fossil record) that the superfamily originated in Africa.[6] Similarly, while coral snakes comprise 16 species in Asia and over 65 species in the Americas, the fact that none of the American clades are basal implies that the group's ancestry was in the Old World.[7]
Notes
- ↑ Meaning the lowest taxonomic ranks of the respective clades; see Amborella example.
- ↑ New Caledonia is viewed as a refugium; i.e., in this case the geographic location of the basal clade is not thought to provide evidence for the locale in which angiosperms originated.
- ↑ This conclusion is consistent with the fact that Didelphimorphia is the basal order within Marsupialia; i.e., extant marsupials as a whole also appear to have originated in South America.[4]
References
- ↑ Essig, F. B. (2014-07-01). "What's so primitive about Amborella?". Botany Professor blog. Retrieved 2014-10-04.
- ↑ Krell, Frank T.; Cranston, Peter S. (2004). "Which side of the tree is more basal?". Systematic Entomology 29: 279–281. doi:10.1111/j.0307-6970.2004.00262.x.
- ↑ Jenner, Ronald A (2006). "Unburdening evo-devo: ancestral attractions, model organisms, and basal baloney". Development Genes and Evolution 216: 385–394. doi:10.1007/s00427-006-0084-5.
- ↑ 4.0 4.1 Nilsson, M. A.; Churakov, G.; Sommer, M.; Van Tran, N.; Zemann, A.; Brosius, J.; Schmitz, J. (2010-07-27). Penny, David, ed. "Tracking Marsupial Evolution Using Archaic Genomic Retroposon Insertions". PLoS Biology (Public Library of Science) 8 (7): e1000436. doi:10.1371/journal.pbio.1000436. PMC 2910653. PMID 20668664.
- ↑ Teeling, E. C.; Springer, M.; Madsen, O.; Bates, P.; O'Brien, S.; Murphy, W. (2005-01-28). "A Molecular Phylogeny for Bats Illuminates Biogeography and the Fossil Record". Science 307 (5709): 580–584. doi:10.1126/science.1105113. PMID 15681385.
- ↑ Gunnell, G. F.; Simmons, N. B.; Seiffert, E. R. (2014-02-04). "New Myzopodidae (Chiroptera) from the Late Paleogene of Egypt: Emended Family Diagnosis and Biogeographic Origins of Noctilionoidea". PLoS ONE 9 (2): e86712. doi:10.1371/journal.pone.0086712. Retrieved 2014-02-05.
- ↑ Slowinski, J. B.; Boundy, J.; Lawson, R. (June 2001). "The Phylogenetic Relationships of Asian Coral Snakes (Elapidae: Calliophis and Maticora) Based on Morphological and Molecular Characters". Herpetologica 57 (2): 233–245. JSTOR 3893186.
- Budd, Graham E.; Jensen, Sören (2000). "A critical reappraisal of the fossil record of the bilaterian phyla". Biological Reviews (Cambridge University Press) 75 (2): 253–95. doi:10.1111/j.1469-185X.1999.tb00046.x. PMID 10881389.
- "Interpreting the Tree Diagram or List of Subgroups on a Tree of Life Page". Tree of Life web project.