Site-specific recombination
Site-specific recombination, also known as conservative site-specific recombination, is a type of genetic recombination in which DNA strand exchange takes place between segments possessing only a limited degree of sequence homology.[1][2][3] Site-specific recombinases (SSRs) perform rearrangements of DNA segments by recognizing and binding to short DNA sequences (sites), at which they cleave the DNA backbone, exchange the two DNA helices involved and rejoin the DNA strands. While in some site-specific recombination systems just a recombinase enzyme and the recombination sites is enough to perform all these reactions, in other systems a number of accessory proteins and/or accessory sites are also needed. Multiple genome modification strategies, among these recombinase-mediated cassette exchange (RMCE), an advanced approach for the targeted introduction of transcription units into predetermined genomic loci, rely on the capacities of SSRs.
Site-specific recombination systems are highly specific, fast and efficient, even when faced with complex eukaryotic genomes.[4] They are employed in a variety of cellular processes, including bacterial genome replication, differentiation and pathogenesis, and movement of mobile genetic elements (Nash 1996). For the same reasons, they present a potential basis for the development of genetic engineering tools.[5]
Recombination sites are typically between 30 and 200 nucleotides in length and consist of two motifs with a partial inverted-repeat symmetry, to which the recombinase binds, and which flank a central crossover sequence at which the recombination takes place. The pairs of sites between which the recombination occurs are usually identical, but there are exceptions (e.g. attP and attB of λ integrase, see lambda phage).[6]
Classification: tyrosine- vs. serine- recombinases

Based on amino acid sequence homology and mechanistic relatedness most site-specific recombinases are grouped into one of two families: the tyrosine recombinase family or the serine recombinase family. The names stem from the conserved nucleophilic amino acid residue that they use to attack the DNA and which becomes covalently linked to it during strand exchange. Early members of the serine recombinase family were known as resolvase/invertases, while the founding member of the tyrosine recombinases, lambda- integrase, using attP/B recognition sites) differs from the now well known enzymes such as Cre (from the P1 phage) and FLP (from yeast S. cerevisiae) while famous serine recombinases include enzymes such as: gamma-delta resolvase (from the Tn1000 transposon), Tn3 resolvase (from the Tn3 transposon) and φC31 integrase (from the φC31 phage).[7]
Although the individual members of the two recombinase families can perform reactions with same practical outcomes, the two families are unrelated to each other, having different protein structures and reaction mechanisms. Unlike tyrosine recombinases, serine recombinases are highly modular as was first hinted by biochemical studies,[8] and later shown by crystallographic structures.[9][10] Knowledge of these protein structures could prove useful when attempting to reengineer recombinase proteins as tools for genetic manipulation.
Mechanism
Recombination between two DNA sites begins by the recognition and binding of these sites by the recombinase protein. This is followed by synapsis, i.e. bringing the sites together to form the synaptic complex. It is within this synaptic complex that the strand exchange takes place, as the DNA is cleaved and rejoined by controlled transesterification reactions. During strand exchange, the DNA cut at fixed points within the crossover region of the site releases a deoxyribose hydroxyl group, while the recombinase protein forms a transient covalent bond to a DNA backbone phosphate. This phosphodiester bond between the hydroxyl group of the nucleophilic serine or tyrosine residue conserves the energy that was expended in cleaving the DNA. Energy stored in this bond is subsequently used for the rejoining of the DNA to the corresponding deoxyribose hydroxyl group on the other site. The entire process therefore goes through without the need for external energy-rich cofactors such as ATP.
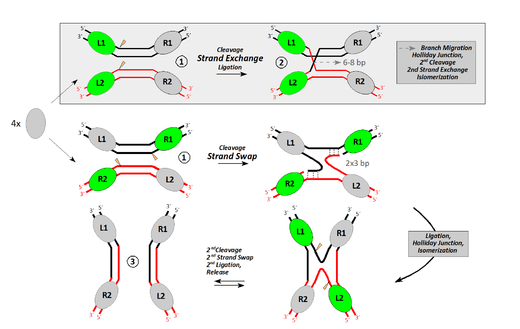
Although the basic chemical reaction is the same for both tyrosine and serine recombinases, there are marked differences. [11] Tyrosine recombinases, such as Cre or Flp, cleave one DNA strand at a time at points that are staggered by 6-8bp, linking the 3’ end of the strand to the hydroxyl group of the tyrosine nucleophile (Fig. 1).[12] Strand exchange then proceeds via a crossed strand intermediate analogous to the Holliday junction in which only one pair of strands has been exchanged.[13][14]
The mechanism and control of serine recombinases is much less well understood. This group of enzymes was only discovered in the mid-1990s and is still relatively small. The now classical members gamma-delta and Tn3 resolvase, but also new additions like φC31-, Bxb1-, and R4 integrases, cut all four DNA strands simultaneously at points that are staggered by 2bp (Fig. 2).[15] During cleavage, a protein-DNA bond is formed via a transesterification reaction in which a phosphodiester bond is replaced by a phosphoserine bond between a 5’ phosphate at the cleavage site and the hydroxyl group of the conserved serine residue (S10 in resolvase).[16][17]
It is still not entirely clear how the strand exchange occurs after the DNA has been cleaved. However, it has been shown that the strands are exchanged while covalently linked to the protein, with a resulting net rotation of 180°.[18][19] The most quoted (but not the only) model accounting for these facts is the "subunit rotation model" (Fig. 2).[11][20] Independent of the model, DNA duplexes are situated outside of the protein complex, and large movement of the protein is needed to achieve the strand exchange. In this case the recombination sites are slightly asymmetric, which allows the enzyme to tell apart the left and right ends of the site. When generating products, left ends are always joined to the right ends of their partner sites, and vice versa. This causes different recombination hybrid sites to be reconstituted in the recombination products. Joining of left ends to left or right to right is avoided due to the asymmetric “overlap” sequence between the staggered points of top and bottom strand exchange, which is in stark contrast to the mechanism employed by tyrosine recombinases.[11]


The reaction catalysed, for instance, by Cre-recombinase may lead to excision of the DNA segment flanked by the two sites (Fig. 3A), but may also lead to integration or inversion of the orientation of the flanked DNA segment (Fig. 3B). What the outcome of the reaction will be is dictated mainly by the relative location and the orientation of sites that are to be recombined, but also by the innate specificity of the site-specific system in question. Excisions and inversions occur if the recombination takes place between two sites that are found on the same molecule (intramolecular recombination), and if the sites are in the same (direct repeat) or in an opposite orientation (inverted repeat), respectively. Insertions, on the other hand, take place if the recombination occurs on sites that are situated on two different DNA molecules (intermolecular recombination), provided that at least one of these molecules is circular. Most site-specific systems are highly specialised, catalysing only one of these different types of reaction, and have evolved to ignore the sites that are in the ‘wrong’ orientation.
See also
- Site-specific recombinase technology
- Recombinase-mediated cassette exchange
- Cre recombinase
- Cre-Lox recombination
- FLP-FRT recombination
- Genetic recombination
- Homologous recombination
References
- ↑ Bode, J; Schlake, T; Iber, M; Schuebeler, D; Seibler, J; Snezhkov, E; Nikolaev, L (2000). "The transgeneticist's toolbox: novel methods for the targeted modification of eukaryotic genomes". Biol. Chem. 381 (9–10): 801–813. doi:10.1515/BC.2000.103. PMID 11076013.
- ↑ Kolb, A.F. (2002). "Genome Engineering Using Site-Specific Recombinases". Cloning & Stem Cells 4 (1): 65–80. doi:10.1089/153623002753632066. PMID 12006158.
- ↑ Coates, C.J.; Kaminski, JM; Summers, JB; Segal, DJ; Miller, AD; Kolb, AF (2005). "Site-directed genome modification: derivatives of DNA-modifying enzymes as targeting tools". Trends in Biotechnology 23 (8): 407–19. doi:10.1016/j.tibtech.2005.06.009. PMID 15993503.
- ↑ Sauer, B. (1998). "Inducible Gene Targeting in Mice Using the Cre/loxSystem". Methods 14 (4): 381–92. doi:10.1006/meth.1998.0593. PMID 9608509.
- ↑ Akopian, A.; Stark, W.M. (2005). "Site-Specific DNA Recombinases as Instruments for Genomic Surgery". Advances in Genetics. Advances in Genetics 55: 1. doi:10.1016/S0065-2660(05)55001-6. ISBN 978-0-12-017655-7.
- ↑ Landy, A. (1989). "Dynamic, Structural, and Regulatory Aspects of lambda Site-Specific Recombination". Annual Reviews in Biochemistry 58 (1): 913–41. doi:10.1146/annurev.bi.58.070189.004405. PMID 2528323.
- ↑ Stark, W.M.; Boocock, M.R. (1995). "Topological selectivity in site-specific recombination". Mobile Genetic Elements. Oxford University Press. pp. 101–29.
- ↑ Abdel-Meguid, S.S.; Grindley, N.D.; Templeton, N.S.; Steitz, T.A. (April 1984). "Cleavage of the site-specific recombination protein gamma delta resolvase: the smaller of two fragments binds DNA specifically". Proc. Natl. Acad. Sci. U.S.A. 81 (7): 2001–5. doi:10.1073/pnas.81.7.2001. PMC 345424. PMID 6326096.
- ↑ Yang, W.; Steitz, T.A. (1995). "Crystal structure of the site-specific recombinase gamma delta resolvase complexed with a 34 bp cleavage site". Cell 82 (2): 193–207. doi:10.1016/0092-8674(95)90307-0. PMID 7628011.
- ↑ Li, W.; Kamtekar, S; Xiong, Y; Sarkis, GJ; Grindley, ND; Steitz, TA (2005). "Structure of a Synaptic Gamma Delta Resolvase Tetramer Covalently Linked to Two Cleaved DNAs". Science 309 (5738): 1210–5. doi:10.1126/science.1112064. PMID 15994378.
- ↑ 11.0 11.1 11.2 Turan, S. , Bode, J., 2011. Review: Site-specific recombinases: from tag-and-target- to tag-and-exchange-based genomic modifications. FASEB J. 25, 4088-4107, http://dx.doi.org/10.1096/fj.11-186940 ; http://www.ncbi.nlm.nih.gov/pubmed/21891781.
- ↑ Van Duyne, G.D. (2002). "A structural view of tyrosine recombinase site-specific recombination". Mobile DNA II. ASM Press. pp. 93–117.
- ↑ Holliday, R. (1964). "A mechanism for gene conversion in fungi". Genetics Research 5 (2): 282–304. doi:10.1017/S0016672300001233.
- ↑ Grainge, I.; Jayaram, M. (1999). "The integrase family of recombinases: organization and function of the active site". Molecular Microbiology 33 (3): 449–56. doi:10.1046/j.1365-2958.1999.01493.x. PMID 10577069.
- ↑ Stark, W.M.; Boocock, M.R.; Sherratt, DJ (1992). "Catalysis by site-specific recombinases". Trends in Genetics 8 (12): 432–9. doi:10.1016/0168-9525(92)90327-Z. PMID 1337225.
- ↑ Reed, R.R.; Grindley, N.D. (1981). "Transposon-mediated site-specific recombination in vitro: DNA cleavage and protein-DNA linkage at the recombination site". Cell 25 (3): 721–8. doi:10.1016/0092-8674(81)90179-3. PMID 6269756.
- ↑ Reed, R.R.; Moser, C.D. (1984). "Resolvase-mediated recombination intermediates contain a serine residue covalently linked to DNA". Cold Spring Harb Symp Quant Biol 49: 245–9. PMID 6099239.
- ↑ Stark, M.W.; Sherratt, DJ; Boocock, MR (1989). "Site-specific recombination by Tn 3 resolvase: topological changes in the forward and reverse reactions". Cell 58 (4): 779–90. doi:10.1016/0092-8674(89)90111-6. PMID 2548736.
- ↑ Stark, W.M.; Boocock, M.R. (1994). "The Linkage Change of a Knotting Reaction Catalysed by Tn3 Resolvase". Journal of Molecular Biology 239 (1): 25–36. doi:10.1006/jmbi.1994.1348. PMID 8196046.
- ↑ Sarkis, G.J; Murley, LL; Leschziner, AE; Boocock, MR; Stark, WM; Grindley, ND (2001). "A model for the gamma-delta resolvase synaptic complex". Molecular Cell 8 (3): 623–31. doi:10.1016/S1097-2765(01)00334-3. PMID 11583624.