Ribonuclease A
| Pancreatic ribonuclease | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Structure of RNase A | |||||||||
| Identifiers | |||||||||
| EC number | 3.1.27.5 | ||||||||
| CAS number | 9001-99-4 | ||||||||
| Databases | |||||||||
| IntEnz | IntEnz view | ||||||||
| BRENDA | BRENDA entry | ||||||||
| ExPASy | NiceZyme view | ||||||||
| KEGG | KEGG entry | ||||||||
| MetaCyc | metabolic pathway | ||||||||
| PRIAM | profile | ||||||||
| PDB structures | RCSB PDB PDBe PDBsum | ||||||||
| Gene Ontology | AmiGO / EGO | ||||||||
| |||||||||

Ribonuclease A (RNase A) is a pancreatic ribonuclease that cleaves single-stranded RNA. Bovine pancreatic RNase A is one of the classic model systems of protein science.[1]
History
The importance of bovine pancreatic RNase A was secured when the Armour & Co. (of hot dog fame) purified a kilogram of it, and gave 10 mg samples away free to any interested scientists.[2] The ability to have a single lot of purified enzyme instantly made RNase a predominant model system for protein studies.
RNase A was the model protein used to work out many spectroscopic methods for assaying protein structure, including absorbance, circular dichroism/optical rotary dispersion, Raman, EPR and NMR spectroscopy. RNase A was also the first model protein for the development of several chemical structural methods, such as limited proteolysis of disordered segments, chemical modification of exposed side chains, and antigenic recognition.
RNase A was the first enzyme for which a correct catalytic mechanism was proposed, even before its structure was known.[3]
Ribonuclease-S, which is RNase A that has been treated with subtilisin, was the third protein to have its structure solved, in 1967.[4]
Studies of the oxidative folding of RNase A led Chris Anfinsen to enunciate the thermodynamic hypothesis of protein folding, which states that the folded form of a protein represents the minimum of its free energy.
RNase A was the first protein for showing the effects of non-native isomers of X-Pro peptide bonds in protein folding.
RNase A was the first protein to be studied by multiple sequence alignment and by comparing the properties of evolutionarily related proteins.
Structure and properties

RNase A is a relatively small protein (124 residues, ~13.7 kDa). It can be characterized as a two-layer 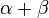 protein that is folded in half to resemble a taco, with a deep cleft for binding the RNA substrate. The first layer is composed of three alpha helices (residues 3-13, 24-34 and 50-60) from the N-terminal half of the protein. The second layer consist of three β-hairpins (residues 61-74, 79-104 and 105-124 from the C-terminal half) arranged in two β-sheets. The hairpins 61-74 and 105-124 form a four-stranded, antiparallel β-sheet that lies on helix 3 (residues 50-60). The longest β-hairpin 79-104 mates with a short β-strand (residues 42-45) to form a three-stranded, antiparallel β-sheet that lies on helix 2 (residues 24-34).
protein that is folded in half to resemble a taco, with a deep cleft for binding the RNA substrate. The first layer is composed of three alpha helices (residues 3-13, 24-34 and 50-60) from the N-terminal half of the protein. The second layer consist of three β-hairpins (residues 61-74, 79-104 and 105-124 from the C-terminal half) arranged in two β-sheets. The hairpins 61-74 and 105-124 form a four-stranded, antiparallel β-sheet that lies on helix 3 (residues 50-60). The longest β-hairpin 79-104 mates with a short β-strand (residues 42-45) to form a three-stranded, antiparallel β-sheet that lies on helix 2 (residues 24-34).
RNase A has four disulfide bonds in its native state: Cys26-Cys84, Cys58-110, Cys40-95 and Cys65-72. The first two (26-84 and 58-110) are essential for conformational folding; each joins an alpha helix of the first layer to a beta sheet of the second layer, forming a small hydrophobic core in its vicinity. The latter two disulfide bonds (40-95 and 65-72) are less essential for folding; either one can be reduced (but not both) without affecting the native structure under physiological conditions. These disulfide bonds connect loop segments and are relatively exposed to solvent. Interestingly, the 65-72 disulfide bond has an extraordinarily high propensity to form, significantly more than would be expected from its loop entropy, both as a peptide and in the full-length protein. This suggests that the 61-74 β-hairpin has a high propensity to fold conformationally.
RNase A is a basic protein (pI = 9.63); its many positive charges are consistent with its binding to RNA (a poly-anion). More generally, RNase A is unusually polar or, rather, unusually lacking in hydrophobic groups, especially aliphatic ones. This may account for its need of four disulfide bonds to stabilize its structure. The low hydrophobic content may also serve to reduce the physical repulsion between highly charged groups (its own and those of its substrate RNA) and regions of low dielectric constant (the nonpolar residues).
The N-terminal α-helix of RNase A (residues 3-13) is connected to the rest of RNase A by a flexible linker (residues 16-23). As shown by F. M. Richards, this linker may be cleaved by subtilisin between residues 20 and 21 without causing the N-terminal helix to dissociate from the rest of RNase A. The peptide-protein complex is called RNase S, the peptide (residues 1-20) is called the S-peptide and the remainder (residues 21-124) is called the S-protein. The dissociation constant of the S-peptide for the S-protein is roughly 30 pM; this tight binding can be exploited for protein purification by attaching the S-peptide to the protein of interest and passing a mixture over an affinity column with bound S-protein. [A smaller C-peptide (residues 1-13) also works.] The RNase S model system has also been used for studying protein folding by coupling folding and association. The S-peptide was the first peptide from a native protein shown to have (flickering) secondary structure in isolation (by Klee and Brown in 1967).
RNase A cleaves specifically after pyrimidine nucleotides.[5] Cleavage takes place in two steps: first, the 3’,5’-phosphodiester bond is cleaved to generate a 2’,3’-cyclic phosphodiester intermediate; second, the cyclic phosphodiester is hydrolyzed to a 3’-monophosphate.[6] It can be inhibited by ribonuclease inhibitor protein, by heavy metal ions, and by uridine-vanadate complexes.[6]
Enzymatic mechanism
The positive charges of RNase A lie mainly in a deep cleft between two lobes. The RNA substrate lies in this cleft and is cleaved by two catalytic histidine residues, His12 and His119, to form a 2',3'-cyclic phosphate intermediate that is stabilized by nearby Lys41.
Enzyme regulation
This protein may use the morpheein model of allosteric regulation.[7]
Anti-cancer effects
RNase A, and to a greater extent its oligomers and some homologs (such as onconase from frogs), have cytotoxic and cytostatic effects, particularly on cancer cells.[8] This attribute has led to the development of onconase as a cancer therapeutic. As with many protein drugs from a non-human source, the internal use of non-human ribonucleases such as onconase is limited by the patient's immune response.
Other biological effects
Ribonuclease is also related to angiogenin, which is involved in blood vessel development.
See also
References
- ↑ Raines, R. T. (1998). "Ribonuclease A". Chem. Rev. 98 (3): 1045–1066. PMID 11848924.
- ↑ Richards FM (November 1972). "The 1972 nobel prize for chemistry". Science 178 (4060): 492–3. doi:10.1126/science.178.4060.492. PMID 17754377.
- ↑ Cuchillo, C. M.; Nogués, M. V.; Raines, R. T. (2011). "Bovine pancreatic ribonuclease: Fifty years of the first enzymatic reaction mechanism". Biochemistry 50 (37): 7835–7841. doi:10.1021/bi201075b. PMC 3172371. PMID 21838247.
- ↑ Wyckoff HW, Hardman KD, Allewell NM, Inagami T, Johnson LN, Richards FM (September 1967). "The structure of ribonuclease-S at 3.5 Å resolution". J. Biol. Chem. 242 (17): 3984–8. PMID 6037556.
- ↑ Volkin E, Cohn WE (December 1953). "On the structure of ribonucleic acids. II. The products of ribonuclease action". J. Biol. Chem. 205 (2): 767–82. PMID 13129256.
- ↑ 6.0 6.1 Krystal Worthington. "Ribonuclease - Worthington Enzyme Manual". Retrieved 2011-09-26.
- ↑ T. Selwood and E. K. Jaffe. (2011). "Dynamic dissociating homo-oligomers and the control of protein function.". Arch. Biochem. Biophys. 519 (2): 131–43. doi:10.1016/j.abb.2011.11.020. PMC 3298769. PMID 22182754.
- ↑ Lee, J. E.; Raines, R. T. (2008). "Ribonucleases as novel chemotherapeutics: The ranpirnase example". BioDrugs 22 (1): 53–58. PMC 2802594. PMID 18215091.
Further reading
- Kartha, G., Bello, J., Harker, D. (1967). Tertiary Structure of Ribonuclease. Boston: Nature. ISBN 0-12-588945-3.
- Scheraga HA, Wedemeyer WJ, Welker E (2001). "Bovine pancreatic ribonuclease A: oxidative and conformational folding studies". Meth. Enzymol. 341: 189–221. doi:10.1016/S0076-6879(01)41153-0. PMID 11582778.
External links
- Ribonuclease A at the US National Library of Medicine Medical Subject Headings (MeSH)
| ||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||