Paxillus involutus
| ||||||||||||||||||||||||||||||||||||||||||||||

Paxillus involutus, commonly known as the brown roll-rim, common roll-rim, or poison pax, is a basidiomycete fungus widely distributed across the Northern Hemisphere. It has been unintentionally introduced to Australia, New Zealand, and South America, where it has likely been transported in soil with European trees. Various shades of brown in colour, the fruit body grows up to 6 cm (2.4 in) high and has a funnel-shaped cap up to 12 cm (5 in) wide with a distinctive inrolled rim and decurrent gills that may be pore-like close to the stipe. Although it has gills, it is more closely related to the pored boletes than to typical gilled mushrooms. It was first described by Pierre Bulliard in 1785, and was given its current binomial name by Elias Magnus Fries in 1838. Genetic testing suggests that Paxillus involutus may be a species complex rather than a single species.
A common mushroom of deciduous and coniferous woods and grassy areas in late summer and autumn, Paxillus involutus forms ectomycorrhizal relationships with a broad range of tree species. These benefit from the symbiosis as the fungus reduces their intake of heavy metals and increases resistance to pathogens such as Fusarium oxysporum. Previously considered edible and eaten widely in Eastern and Central Europe, it has since been found to be dangerously poisonous, responsible for the death of German mycologist Julius Schäffer in 1944. It had been recognised as causing gastric upsets when eaten raw, but was more recently found to cause potentially fatal autoimmune hemolysis, even in those who had consumed the mushroom for years without any other ill effects. An antigen in the mushroom triggers the immune system to attack red blood cells. Serious and potentially fatal complications include acute renal failure, shock, acute respiratory failure, and disseminated intravascular coagulation.
Taxonomy and naming
.jpg)
The brown roll-rim was described by French mycologist Pierre Bulliard in 1785 as Agaricus contiguus,[3] although the 1786 combination Agaricus involutus of August Batsch[4] is taken as the first valid description.[1] James Bolton published a description of what he called Agaricus adscendibus in 1788;[5] the taxonomical authority Index Fungorum considers this to be synonymous with P. involutus.[2] Additional synonyms include Omphalia involuta described by Samuel Frederick Gray in 1821,[6] and Rhymovis involuta, published by Gottlob Ludwig Rabenhorst in 1844.[7] The species gained its current binomial name in 1838 when the 'father of mycology', Swedish naturalist Elias Magnus Fries erected the genus Paxillus, and set it as the type species.[8] The starting date of fungal taxonomy had been set as January 1, 1821, to coincide with the date of Fries' works, which meant that names coined earlier than this date required sanction by Fries (indicated in the name by a colon) to be considered valid. It was thus written Paxillus involutus (Batsch:Fr.) Fr. However, a 1987 revision of the International Code of Botanical Nomenclature set the starting date at May 1, 1753, the date of publication of Linnaeus' seminal work, the Species Plantarum.[9] Hence the name no longer requires the ratification of Fries' authority.
The genus was later placed in a new family, Paxillaceae, by French mycologist René Maire who held it to be related to both agarics and boletes.[10] Although it has gills rather than pores, it has long been recognised as belonging to the pored mushrooms of the order Boletales rather than the traditional agarics.[11] The generic name is derived from the Latin for 'peg' or 'plug', and the specific epithet involutus, 'inrolled', refers to the cap margin.[11] Common names include the naked brimcap,[12] poison paxillus,[13] inrolled pax,[14] poison pax, common roll-rim, brown roll-rim,[15] and brown chanterelle.[16] Gray called it the "involved navel-stool" in his 1821 compendium of British flora.[6]
Studies of the ecology and genetics of Paxillus involutus indicate that it may form a complex of multiple similar-looking species.[17][18] In a field study near Uppsala, Sweden, conducted from 1981 to 1983, mycologist Nils Fries found that there were three populations of P. involutus unable to breed with each other. One was found under conifers and mixed woodlands, while the other two were found in parklands, associated with nearby birch trees. He found that the first group tended to produce single isolated fruit bodies which had a thinner stipe and cap which was less inrolled at the margins, while the fruit bodies of the other two populations tended to appear in groups, and have thicker stipes, and caps with more inrolled and sometimes undulating margins. However, there were only general tendencies and he was unable to detect any consistent macroscopic or microscopic features that firmly differentiate them.[19] A molecular study comparing the DNA sequences of specimens of Paxillus involutus collected from various habitats in Bavaria found that those collected from parks and gardens showed a close relationship with the North American species P. vernalis, while those from forests were allied with P. filamentosus. The authors suggested the park populations may have been introduced from North America.[20] A multi-gene analysis of European isolates showed that P. involutus sensu lato (in the loose sense) could be separated into four distinct, genetically isolated lineages corresponding to P. obscurosporus, P. involutus sensu stricto (in the strict sense), P. validus, and a fourth species that has not yet been identified.[18] Changes in host range have occurred frequently and independently among strains within this species complex.[21]
Description

Resembling a brown wooden top, the epigeous (aboveground) fruit body may be up to 6 cm (2.4 in) high.[11] The cap, initially convex then more funnel-shaped (infundibuliform) with a depressed centre and rolled rim (hence the common name), may be reddish-, yellowish- or olive-brown in colour and typically 4–12 cm (1.6–5 in) wide;[22] the cap diameter does not get larger than 15 cm (5.9 in).[23] The cap surface is initially downy and later smooth, becoming sticky when wet. The cap and cap margin initially serve to protect the gills of young fruit bodies: this is termed pilangiocarpic development.[24] The narrow brownish yellow gills are decurrent and forked, and can be peeled easily from the flesh (as is the case with the pores of boletes). Gills further down toward the stipe become more irregular and anastomose, and can even resemble the pores of bolete-type fungi. The fungus darkens when bruised and older specimens may have darkish patches. The juicy yellowish flesh has a mild to faintly sour or sharp odor and taste, and has been described as well-flavored upon cooking.[22][25] Of similar colour to the cap, the short stipe can be crooked and tapers toward the base.[26]
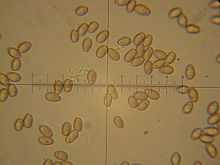
The spore print is brown, and the dimensions of the ellipsoid (oval-shaped) spores are 7.5–9 by 5–6 μm. The hymenium has cystidia both on the gill edge and face (cheilo- and pleurocystidia respectively), which are slender and filament-like, typically measuring 40–65 by 8–10.5 μm.[27]
Similar species
The brownish colour and funnel-like shape of P. involutus can lead to its confusion with several species of Lactarius, many of which have some degree of toxicity themselves.[28] However, the lack of a milky exudate distinguishes it from any milk cap.[29] One of the more similar is L. turpis, which presents a darker olive colouration.[25] The related North American Paxillus vernalis has a darker spore print, thicker stipe and is found under aspen,[13] whereas the closer relative P. filamentosus is more similar in appearance to P. involutus. A rare species that grows only in association with alder, P. filamentosus can be distinguished from it by the pressed-down scales on the cap surface that point towards the cap margin, a light yellow flesh that bruises only slightly brown, and deep yellow-ochre gills that do not change colour upon injury[27]
The most similar species are two once thought to be part of P. involutus in Europe. Paxillus obscurosporus has larger fruit bodies than P. involutus, with caps up to 40 cm (16 in) wide whose margins tend to unroll and flatten with age, and a layer of cream-coloured mycelia covering the base of its tapered stipe. P. validus, also known only from Europe, has caps up to 20 cm (7.9 in) wide with a stipe that is more or less equal in width throughout its length. Found under broadleaved trees in parks, it can be reliably distinguished from P. involutus (and other Paxillus species) by the presence of crystals up to 2.5 μm long in the rhizomorphs, as the crystals found in rhizomorphs of other Paxillus species do not exceed 0.5 μm long.[23]
Ecology, distribution and habitat
Paxillus involutus forms ectomycorrhizal relationships with a number of coniferous and deciduous tree species. Because the fungus has somewhat unspecialised nutrient requirements and a relatively broad host specificity, it has been frequently used in research and seedling inoculation programs.[30] There is evidence of the benefit to trees of this arrangement: in one experiment where P. involutus was cultivated on the root exudate of red pine (Pinus resinosa), the root showed markedly increased resistance to pathogenic strains of the ubiquitous soil fungus Fusarium oxysporum.[31] Seedlings inoculated with P. involutus also showed increased resistance to Fusarium.[32] Thus P. involutus may be producing antifungal compounds which protect the host plants from root rot.[33] Paxillus involutus also decreases the uptake of certain toxic elements, acting as a buffer against heavy metal toxicity in the host plant. For example, the fungus decreased the toxicity of cadmium and zinc to Scots pine (Pinus sylvestris) seedlings: even though cadmium itself inhibits ectomycorrhiza formation in seedlings, colonization with P. involutus decreases cadmium and zinc transport to the plant shoots and alters the ratio of zinc transported to the roots and shoots, causing more cadmium to be retained in the roots of the seedlings rather than distributed through its entire metabolism.[34] Evidence suggests that the mechanism for this detoxification involves the cadmium binding to the fungal cell walls, as well as accumulating in the vacuolar compartments.[35] Further, ectomycorrhizal hyphae exposed to copper[36] or cadmium drastically increase production of a metallothionein—a low molecular weight protein that binds metals.[37][38]
The presence of Paxillus involutus is related to much reduced numbers of bacteria associated with the roots of Pinus sylvestris. Instead bacteria are found on the external mycelium.[39] The types of bacteria change as well; a Finnish study published in 1997 found that bacterial communities under P. sylvestris without mycorrhizae metabolised organic and amino acids, while communities among P. involutus metabolised the sugar fructose.[40] Paxillus involutus benefits from the presence of some species of bacteria in the soil it grows in. As the fungus grows it excretes polyphenols, waste products that are toxic to itself and impede its growth, but these compounds are metabolised by some bacteria, resulting in increased fungal growth. Bacteria also produce certain compounds such as citric and malic acid, which stimulate P. involutus.[41]

Highly abundant,[29] the brown roll-rim is found across the Northern Hemisphere, Europe and Asia, with records from India,[42] China,[43] Japan, Iran,[44] and Turkey's eastern Anatolia.[45] It is equally widely distributed across northern North America,[28] extending north to Alaska, where it has been collected from tundra near Coldfoot in the interior of the state.[46] In southwestern Greenland, P. involutus has been recorded under the birch species Betula nana, B. pubescens and B. glandulosa.[47] The mushroom is more common in coniferous woods in Europe, but is also closely associated with birch (Betula pendula). Within woodland, it prefers wet places or boggy ground, and avoids calcareous (chalky) soils. It has been noted to grow alongside Boletus badius in Europe,[22] and Leccinum scabrum and Lactarius plumbeus in the Pacific Northwest region of North America.[48] There it is found in both deciduous and coniferous woodland, commonly under plantings of white birch (Betula papyrifera) in urban areas.[28] It is one of a small number of fungal species which thrive in Pinus radiata plantations planted outside their natural range.[49] A study of polluted Scots pine forest around Oulu in northern Finland found that P. involutus became more abundant in more polluted areas while other species declined. Emissions from pulp mills, fertiliser, heating and traffic were responsible for the pollution, which was measured by sulfur levels in the pine needles.[50]
Paxillus involutus can be found growing on lawns and old meadows throughout its distribution. Fruit bodies are generally terrestrial, though they may be found on woody material around tree stumps.[28] They generally appear in autumn and late summer.[22] In California, David Arora discerned a larger form associated with oak and pine which appears in late autumn and winter, as well as the typical form that is associated with birch plantings and appears in autumn.[14] Several species of flies and beetles have been recorded using the fruit bodies to rear their young.[51] The mushroom can be infected by Hypomyces chrysospermus, or bolete eater, a mould species that parasitises Boletales members.[52] Infection results in the appearance of a whitish powder that first manifests on the pores, then spreads over the surface of the mushroom, becoming golden yellow to reddish-brown in maturity.[53]
Australian mycologist John Burton Cleland noted it occurring under larch (Larix), oak, pine, birch and other introduced trees in South Australia in 1934,[54] and it has subsequently been recorded in New South Wales, Victoria[55] (where it was found near Betula and Populus)[56] and Western Australia. It has been recorded under introduced birch (Betula) and hazel (Corylus) in New Zealand.[57] Mycologist Rolf Singer reported a similar situation in South America, with the species recorded under introduced trees in Chile. It is likely to have been transported to those countries in the soil of imported European trees.[58]
Toxicity

Paxillus involutus was widely eaten in Central and Eastern Europe until World War II, although English guidebooks did not recommend it.[22][29] In Poland, the mushroom was often eaten after pickling or salting.[15] It was known to be a gastrointestinal irritant when ingested raw but had been presumed edible after cooking.[27] Questions were first raised about its toxicity after German mycologist Julius Schäffer died after eating it in October 1944. About an hour after he and his wife ate a meal prepared with the mushrooms, Schäffer developed vomiting, diarrhea, and fever. His condition worsened to the point where he was admitted to hospital the following day and developed renal failure, perishing after 17 days.[29][59]
In the mid-1980s, Swiss physician René Flammer discovered an antigen within the mushroom that stimulates an autoimmune reaction causing the body's immune cells to consider its own red blood cells as foreign and attack them. Despite this, it was not until 1990 that guidebooks firmly warned against eating P. involutus, and one Italian guidebook recommended it as edible in 1998.[60] The relatively rare immunohemolytic syndrome occurs following the repeated ingestion of Paxillus mushrooms.[61] Most commonly it arises when the person has ingested the mushroom for a long period of time, sometimes for many years, and has shown mild gastrointestinal symptoms on previous occasions.[27] The Paxillus syndrome is better classed as an allergic reaction than a toxicological reaction as it is not caused by a genuinely poisonous substance but by the antigen in the mushroom. The antigen is still of unknown structure but it stimulates the formation of IgG antibodies in the blood serum. In the course of subsequent meals, antigen-antibody complexes are formed; these complexes attach to the surface of blood cells and eventually lead to their breakdown.[27]
Poisoning symptoms are rapid in onset, consisting initially of vomiting, diarrhea, abdominal pain, and associated decreased blood volume.[62] Shortly after these initial symptoms appear, hemolysis develops, resulting in reduced urine output, hemoglobin in the urine or outright absence of urine formation, and anemia. Medical laboratory tests consist of testing for the presence of increasing bilirubin and free hemoglobin, and falling haptoglobins. Hemolysis may lead to numerous complications including acute renal failure, shock, acute respiratory failure, and disseminated intravascular coagulation.[27][63][64] These complications can cause significant morbidity with fatalities having been reported.[63]
There is no antidote for poisoning, only supportive treatment consisting of monitoring complete blood count, renal function, blood pressure, and fluid and electrolyte balance[65] and correcting abnormalities. The use of corticosteroids may be a useful adjunct in treatment, as they protect blood cells against hemolysis, thereby reducing complications.[66] Plasmapheresis reduces the circulating immune complexes in the blood which cause the hemolysis, and may be beneficial in improving the outcome.[62][67] Additionally, hemodialysis can be used for patients with compromised renal function or renal failure.[27]
Paxillus involutus also contains agents which appear to damage chromosomes; it is unclear whether these have carcinogenic or mutagenic potential.[68] Two compounds that have been identified are the phenols involutone[69] and involutin; the latter is responsible for the brownish discolouration upon bruising.[16]
See also
References
- ↑ 1.0 1.1 "Paxillus involutus (Batsch) Fr. 1838". MycoBank. International Mycological Association. Retrieved 2011-06-29.
- ↑ 2.0 2.1 "Agaricus adscendibus Bolton". Index Fungorum. CAB International. Retrieved 2011-07-15.
- ↑ Bulliard, J.B.F. (1785). Herbier de la France (in French) 5. Paris, France: Chez l'auteur. pp. 192–240.
- ↑ Batsch, August (1786). Elenchus Fungorum (Discussion of Fungi), Continuatio Prima (in Latin). Halle, Magdeburg, Germany: Apud Joannem J. Gebauer. p. 39.
- ↑ Bolton, James (1788). An History of Fungusses, Growing about Halifax 2. Huddersfield, UK: self-published. p. 55.
- ↑ 6.0 6.1 Gray, Samuel F. (1821). Natural Arrangement of British Plants. London, UK: Baldwin, Cradock, and Joy. p. 611.
- ↑ Rabenhorst, Gottlob Ludwig (1844). Deutschlands Kryptogamenflora (in German) 1 (2 ed.). Leipzig, Germany: E. Kummer. p. 453.
- ↑ Fries, Elias Magnus (1838). Epicrisis Systematis Mycologici (in Latin). Uppsala, Sweden: Typographia Academica. p. 317.
- ↑ Esser, K.; Lemke, P.A. (1994). The Mycota: A Comprehensive Treatise on Fungi as Experimental Systems for Basic and Applied Research. Heidelberg, Germany: Springer. p. 81. ISBN 3-540-66493-9.
- ↑ Maire, René (1902). "Recherches cytologiques et taxonomiques sur les Basidiomycetes". Bulletin trimestriel de la Société Mycologique de France (in French) 18 (supplement): 165.
- ↑ 11.0 11.1 11.2 Nilson, Sven; Persson, Ole (1977). Fungi of Northern Europe 1: Larger Fungi (Excluding Gill-Fungi). Harmondsworth, UK: Penguin. p. 121. ISBN 0-14-063005-8.
- ↑ Ziobro, G. (2012). "Mushroom Toxins" (PDF). In Lampel, Keith A.; Al-Khaldi, Sufian; Cahil, Susan Mary. Bad Bug Book: Foodborne Pathogenic Microorganisms and Natural Toxins Handbook (2nd ed.). USFDA/Center for Food Safety & Applied Nutrition. p. 204.
- ↑ 13.0 13.1 Lincoff, Gary H. (2008). National Audubon Society Field Guide to North American Mushrooms. New York, New York: Alfred A. Knopf. p. 671. ISBN 0-394-51992-2.
- ↑ 14.0 14.1 Arora, David (1986). Mushrooms Demystified: a Comprehensive Guide to the Fleshy Fungi. Berkeley, California: Ten Speed Press. pp. 477–78. ISBN 0-89815-169-4.
- ↑ 15.0 15.1 Benjamin, Denis R. (1995). Mushrooms: Poisons and Panaceas—A Handbook for Naturalists, Mycologists and Physicians. New York, New York: WH Freeman and Company. pp. 382–84. ISBN 0-7167-2600-9.
- ↑ 16.0 16.1 Barceloux, Donald G. (2008). Medical Toxicology of Natural Substances: Foods, Fungi, Medicinal Herbs, Plants, and Venomous Animals. Hoboken, New Jersey: John Wiley and Sons. pp. 312–13. ISBN 0-471-72761-X.
- ↑ Hahn, C.; Agerer, R. (1999). "Studien zum Paxillus involutus Formenkreis" [Studies on the Paxillus involutus complex]. Nova Hedwigia (in German) 69: 241–310.
- ↑ 18.0 18.1 Hedh, Jenny; Samson, Peter; Erland, Susanne; Tunlid, Anders (2008). "Multiple gene genealogies and species recognition in the ectomycorrhizal fungus Paxillus involutus". Mycological Research 112 (8): 965–75. doi:10.1016/j.mycres.2008.01.026. PMID 18554888.
- ↑ Fries, Nils (1985). "Intersterility groups in Paxillus involutus". Mycotaxon 24: 403–10.
- ↑ Jarosch, Margit; Bresinsky, A. (1999). "Speciation and phylogenetic distances within Paxillus s. str. (Basidiomycetes, Boletales)". Plant Biology 1 (6): 701–05. doi:10.1111/j.1438-8677.1999.tb00283.x.
- ↑ Hehd, Jenny; Johannsson, Tomas; Tunlid, Anders (2009). "Variation in host specificity and gene content in strains from genetically isolated lineages of the ectomycorrhizal fungus Paxillus involutus s. lat.". Mycorrhiza 19 (8): 549–58. doi:10.1007/s00572-009-0252-3. PMID 19452174.
- ↑ 22.0 22.1 22.2 22.3 22.4 Haas, Hans (1969). The Young Specialist looks at Fungi. London, UK: Burke. p. 54. ISBN 0-222-79409-7.
- ↑ 23.0 23.1 Henrici, Alick (2004). "A key to Paxillus s.l. in Europe". Field Mycology 5 (3): 87–88. doi:10.1016/S1468-1641(10)60560-9.
- ↑ Chiu, Siu-Wai; Moore, David (1996). Patterns in Fungal Development. Cambridge, UK: Cambridge University Press. p. 207. ISBN 0-521-56047-0.
- ↑ 25.0 25.1 Zeitlmayr, Linus (1976). Wild Mushrooms: An Illustrated Handbook. Hertfordshire, UK: Garden City Press. p. 75. ISBN 0-584-10324-7.
- ↑ Breitenbach, J.; Kränzlin, F. (1991). Fungi of Switzerland 3: Boletes & Agarics, 1st Part. Lucerne, Switzerland: Edition Mykologia. ISBN 3-85604-230-X.
- ↑ 27.0 27.1 27.2 27.3 27.4 27.5 27.6 Bresinsky, Andreas; Besl, Helmut (1990). A Colour Atlas of Poisonous Fungi. London, UK: Wolfe Publishing. pp. 126–29. ISBN 0-7234-1576-5.
- ↑ 28.0 28.1 28.2 28.3 Ammirati, Joseph F.; Traquair, James A.; Horgen, Paul A. (1985). Poisonous Mushrooms of the Northern United States and Canada. Minneapolis, Minnesota: University of Minnesota Press. pp. 78 , 293–95. ISBN 0-8166-1407-5.
- ↑ 29.0 29.1 29.2 29.3 Lamaison, Jean-Louis; Polese, Jean-Marie (2005). The Great Encyclopedia of Mushrooms. Cologne, Germany: Könemann. p. 35. ISBN 3-8331-1239-5.
- ↑ Taylor A.F.S.; Martin, F.; Read, D.J. (2000). "Fungal diversity in ectomycorrhizal communities of Norway Spruce [Picea abies (L.) Karst.] and Beech (Fagus sylvatica L.) along north-south transects in Europe". In Schulze, Ernst-Detlef. Carbon and Nitrogen Cycling in European Forest Ecosystems. Ecological Studies. Berlin, Germany: Springer. pp. 343–65. ISBN 978-3-540-67025-4.
- ↑ Duchesne, Luc C.; Peterson, R.L.; Ellis, Brian E. (1988). "Pine root exudate stimulates the synthesis of antifungal compounds by the ectomycorrhizal fungus Paxillus involutus". New Phytologist 108 (4): 471–76. doi:10.1111/j.1469-8137.1988.tb04188.x. JSTOR 2432861.
- ↑ Duchesne, Luc C.; Peterson, R.L.; Ellis, Brian E. (1988). "Interaction between the ectomycorrhizal fungus Paxillus involutus and Pinus resinosa induces resistance to Fusarium oxysporum". Canadian Journal of Botany 66 (3): 558–62. doi:10.1139/b88-080.
- ↑ Duchesne, Luc C.; Peterson, R.L.; Ellis, Brian E. (1989). "The time-course of disease suppression and antibiosis by the ectomycorrhizal fungus Paxillus involutus". New Phytologist 111 (4): 693–98. doi:10.1111/j.1469-8137.1989.tb02364.x. JSTOR 2556679.
- ↑ Dixon, Robert K. (1988). "Response of ectomycorrhizal Quercus rubra to soil cadmium, nickel and lead". Soil Biology and Biochemistry 20 (4): 555–59. doi:10.1016/0038-0717(88)90072-7.
- ↑ Marschner, Petra; Jentschke, Georg; Godbold, Douglas L. (1998). "Cation exchange capacity and lead sorption in ectomycorrhizal fungi". Plant Soil 205 (1): 93–98. doi:10.1023/A:1004376727051.
- ↑ Blaudez, Damien; Botton, Bernard; Chalot, Michel (2000). "Cadmium uptake and subcellular compartmentation in the ectomycorrhizal fungus Paxillus involutus" (PDF). Microbiology 146 (5): 1109–17. PMID 10832638.
- ↑ Morselt, A.F.W.; Smits, W.T.M.; Limonard, T. (1986). "Histochemical demonstration of heavy metal tolerance in ectomycorrhizal fungi". Plant Soil 96 (3): 417–20. doi:10.1007/BF02375146.
- ↑ Bellion, Marc; Courbot, Mikael; Jacob, Christian; Guinet, Frédéric; Blaudez, Damien; Chalot, Michel (2007). "Metal induction of a Paxillus involutus metallothionein and its heterologous expression in Hebeloma cylindrosporum". New Phytologist 174 (1): 151–58. doi:10.1111/j.1469-8137.2007.01973.x.
- ↑ Nurmiaho-Lassila, E.-L.; Timonen, S.; Haahtela, K.; Sen, R. (1997). "Bacterial colonization patterns of intact Pinus sylvestris mycorrhizospheres in dry pine forest soil: an electron microscopy study". Canadian Journal of Microbiology 43 (11): 1017–35. doi:10.1139/m97-147.
- ↑ Timonen, Sari; Jørgensen, Kirsten S.; Haahtela, Kielo; Sen, Robin (1997). "Bacterial community structure at defined locations of Pinus sylvestris–Suillus bovinus and Pinus sylvestris–Paxillus involutus mycorrhizospheres in dry pine forest humus and nursery peat". Canadian Journal of Microbiology 44 (6): 499–513. doi:10.1139/w98-035.
- ↑ Duponnois, R.; Garbaye, J. (1990). "Some mechanisms involved in growth stimulation of ectomycorrhizal fungi by bacteria". Canadian Journal of Botany 68 (10): 2148–52. doi:10.1139/b90-280.
- ↑ Mukeriji, K.G.; Manoharachary, C. (2010). Taxonomy and Ecology of Indian Fungi. New Dehli, India: I.K. International Publishing House. p. 204. ISBN 978-93-8002-692-3.
- ↑ Zhishu, Bi; Zheng, Guoyang; Taihui, Li (1993). The Macrofungus Flora of China's Guangdong Province (Chinese University Press). New York, New York: Columbia University Press. p. 455. ISBN 962-201-556-5.
- ↑ Asef Shayan, M.R. (2010). قارچهای سمی ایران (Qarch-ha-ye Sammi-ye Iran) [Poisonous mushrooms of Iran] (in Persian). Iran shenasi. p. 214. ISBN 978-964-2725-29-8.
- ↑ Demirel, K.; Uzun, Y.; Kaya, A. (2004). "Some poisonous fungi of east Anatolia" (PDF). Turkish Journal of Botany 28: 215–19.
- ↑ Laursen, Gary A.; Seppelt, Rodney D. (2009). Common Interior Alaska Cryptogams: Fungi, Lichenicolous Fungi, Lichenized Fungi, Slime Molds, Mosses, and Liverworts. College, Alaska: University of Alaska Press. p. 60. ISBN 1-60223-058-7.
- ↑ Knudson, Henning (2006). Arctic and Alpine Mycology 6. Copenhagen, Denmark: Museum Tusculanum Press. p. 12. ISBN 87-635-1277-7.
- ↑ Ammirati, Joseph; Trudell, Steve (2009). Mushrooms of the Pacific Northwest: Timber Press Field Guide (Timber Press Field Guides). Portland, Oregon: Timber Press. p. 184. ISBN 0-88192-935-2.
- ↑ Richardson, David M. (2000). Ecology and Biogeography of Pinus. Cambridge University Press. p. 333. ISBN 0-521-78910-9.
- ↑ Tarvainen, Oili; Markkola, Anna Mari; Strömmer, Rauni (2003). "Diversity of macrofungi and plants in Scots pine forests along an urban pollution gradient". Basic and Applied Ecology 4 (6): 547–56. doi:10.1078/1439-1791-00156.
- ↑ Bruns, Thomas D. (1984). "Insect mycophagy in the Boletales: fungivore diversity and the mushroom habitat". In Blackwell, Meredith; Wheeler, Quentin. Fungus–Insect Relationships: Perspectives in Ecology and Evolution. New York, New York: Columbia University Press. pp. 91–129. ISBN 0-231-05695-8.
- ↑ Sahr, Tobias; Ammer, Hubert; Besl, Helmut; Fischer, Michael (1999). "Infrageneric classification of the boleticolous genus Sepedonium: species delimitation and phylogenetic relationships". Mycologia 91 (6): 935–43. doi:10.2307/3761625. JSTOR 3761625.
- ↑ Kuo, Michael (February 2004). "Hypomyces chrysospermus". MushroomExpert. Retrieved 2011-07-26.
- ↑ Cleland, John Burton (1976). Toadstools and Mushrooms and Other Larger Fungi of South Australia. Adelaide, Australia: South Australian Government Printer. p. 177.
- ↑ Young, A.M. (2004). A Field Guide to the Fungi of Australia. New South Wales, Australia: University of New South Wales Press. p. 169. ISBN 0-86840-742-9.
- ↑ Watling, R.; Hui, L.T. (1999). Australian Boletes – A Preliminary Survey. Edinburgh, Scotland: Royal Botanic Gardens Edinburgh. p. 64. ISBN 1-872291-28-7.
- ↑ McNabb, R.F.R. (1969). "The Paxillaceae of New Zealand". New Zealand Journal of Botany 7 (4): 349–62.
- ↑ Palfner, Götz (2004). "Macrofungi from Chile". self. Retrieved 2011-07-14.
- ↑ Beuchat, Larry R. (1987). Food and Beverage Mycology. New York, New York: Springer. p. 394. ISBN 0-442-21084-1.
- ↑ Marley, Greg (2010). Chanterelle Dreams, Amanita Nightmares: The Love, Lore, and Mystique of Mushrooms. White River Junction, Vermont: Chelsea Green Publishing. pp. 136–40. ISBN 978-1-60358-214-8.
- ↑ Flammer, René (1985). "[Paxillus syndrome: immunohemolysis following repeated mushroom ingestion]". Schweizerische Rundschau fur Medizin Praxis (in German) 74 (37): 997–99. PMID 4059740.
- ↑ 62.0 62.1 Winkelmann, M.; Stangel, W.; Schedel, I.; Grabensee, B. (1986). "Severe hemolysis caused by antibodies against the mushroom Paxillus involutus and its therapy by plasma exchange". Klinische Wochenschrift 64 (19): 935–38. doi:10.1007/BF01728620. PMID 3784443.
- ↑ 63.0 63.1 Winkelmann, M.; Borchard, F.; Stangel, W.; Grabensee, B. (1982). "[Fatal immunohaemolytic anaemia after eating the mushroom Paxillus involutus (author's transl)]". Deutsche Medizinische Wochenschrift (in German) 107 (31–32): 1190–94. doi:10.1055/s-2008-1070100. PMID 7105997.
- ↑ Schmidt, J.; Hartmann, W.; Würstlin, A.; Deicher, H. (1971). "[Acute kidney failure due to immunohemolytic anemia following consumption of the mushroom Paxillus involutus]". Deutsche Medizinische Wochenschrift (in German) 96 (28): 1188–91. doi:10.1055/s-0028-1110104. PMID 5105189.
- ↑ Köppel, C. (1993). "Clinical symptomatology and management of mushroom poisoning". Toxicon 31 (12): 1513–40. doi:10.1016/0041-0101(93)90337-I. PMID 8146866.
- ↑ Olesen, L.L. (1991). "[Poisoning with the brown roll-rim mushroom, Paxillus involutus]". Ugeskrift for Laeger (in Danish) 153 (6): 445. PMID 2000656.
- ↑ Musselius, S.G.; Ryk, A.A.; Lebedev, A.G.; Pakhomova, G.V.; Golikov, P.P.; Davydov, B.V.; Donova, L.V.; Zimina, L.N.; Platonova, G.A.; Selina, I.E.; Skvortsova, A.V. (2002). "[Toxicity of mushrooms Paxillus involutus and Paxillus atrotomentosus]". Anesteziologiia i Reanimatologiia (in Russian) (2): 30–35. PMID 12226995.
- ↑ Gilot-Delhalle, J.; Moutschen, J.; Moutschen-Dahmen, M. (1991). "Chromosome-breaking activity of extracts of the mushroom Paxillus involutus Fries ex Batsch". Experientia 47 (3): 282–84. doi:10.1007/BF01958161. PMID 2009940.
- ↑ Antkowiak, Róza; Antkowiak, Wieslaw Z; Banczyk, Izabela; Mikolajczyk, Lucyna (2003). "A new phenolic metabolite, involutone, isolated from the mushroom Paxillus involutus". Canadian Journal of Chemistry 81 (1): 118–24. doi:10.1139/v02-194.
External links
| Wikimedia Commons has media related to Paxillus involutus. |