Ovule

Ovule means "small egg". In seed plants, the ovule is the structure that gives rise to and contains the female reproductive cells. It consists of three parts: The integument(s) forming its outer layer(s), the nucellus (or remnant of the megasporangium), and the megaspore-derived female gametophyte (or megagametophyte) in its center. The megagametophyte (also called embryo sac in flowering plants) produces an egg cell (or several egg cells in some groups) for fertilization. After fertilization, the ovule develops into a seed.
Location within the plant
In flowering plants, the ovule is typically located inside the portion of the flower called the gynoecium. The ovary of the gynoecium produces one or more ovules and ultimately becomes the fruit wall. Ovules are attached to placenta in the ovary through a stalk-like structure known as a funiculus. Different patterns of ovule attachment, or placentation, can be found among plant species.
In gymnosperms such as conifers, ovules are typically borne on the surface of an ovuliferous (ovule-bearing) scale, usually within an ovulate cone (also called megastrobilus).
In some extinct plants (e.g. Pteridosperms), megasporangia and perhaps ovules were borne on the surface of leaves. In other extinct taxa, a cupule (a modified leaf or part of a leaf) surrounds the ovule (e.g. Caytonia or Glossopteris).
Ovule parts and development
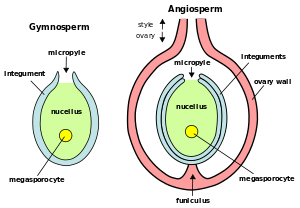

The ovule appears to be a megasporangium with integuments surrounding it. Ovules are initially composed of diploid maternal tissue, which includes a megasporocyte (a cell that will undergo meiosis to produce megaspores). Megaspores remain inside the ovule and divide by mitosis to produce the haploid female gametophyte or megagametophyte, which also remains inside the ovule. The remnants of the megasporangium tissue (the nucellus) surround the megagametophyte. Megagametophytes produce archegonia (lost in some groups such as flowering plants), which produce egg cells. After fertilization, the ovule contains a diploid zygote and then, after cell division begins, an embryo of the next sporophyte generation. In flowering plants, a second sperm nucleus fuses with other nuclei in the megagametophyte forming a typically polyploid (often triploid) endosperm tissue, which serves as nourishment for the young sporophyte.
Integuments, micropyle and chalaza
An integument is a protective cell layer surrounding the ovule. Gymnosperms typically have one integument (unitegmic) while angiosperms typically have two (bitegmic). The evolutionary origin of the inner integument (which is integral to the formation of ovules from megasporangia) has been proposed to be by enclosure of a megasporangium by sterile branches (telomes).[1] Elkinsia, a preovulate taxon, has a lobed structure fused to the lower third of the megasporangium, with the lobes extending upwards in a ring around the megasporangium. This might, through fusion between lobes and between the structure and the megasporangium, have produced an integument.[2]
The origin of the second or outer integument has been an area of active contention for some time. The cupules of some extinct taxa have been suggested as the origin of the outer integument. A few angiosperms produce vascular tissue in the outer integument, the orientation of which suggests that the outer surface is morphologically abaxial. This suggests that cupules of the kind produced by the Caytoniales or Glossopteridales may have evolved into the outer integument of angiosperms.[3]
The integuments develop into the seed coat when the ovule matures after fertilization.
The integuments do not enclose the nucellus completely but retain an opening at the apex referred to as the micropyle. The micropyle opening allows pollen to enter the ovule for fertilization. In gymnosperms (e.g., conifers), pollen is drawn into the ovule on a drop of fluid that exudes out of the micropyle. Subsequently, the micropyle closes. In angiosperms, only a pollen tube enters the micropyle. During germination, the seedling's radicle emerges through the micropyle.
Located opposite from the micropyle is the chalaza where the nucellus is joined to the integuments. Nutrients from the plant travel through the phloem of the vascular system to the funiculus and outer integument and from there apoplastically and symplastically through the chalaza to the nucellus inside the ovule. In chalazogamous plants, the pollen tubes enter the ovule through the chalaza instead of the micropyle opening.
Nucellus, megaspore and perisperm
The nucellus (plural: nucelli) is part of the inner structure of the ovule, forming a layer of diploid (sporophytic) cells immediately inside the integuments. It is structurally and functionally equivalent to the megasporangium. In immature ovules, the nucellus contains a megasporocyte (megaspore mother cell), which undergoes sporogenesis via meiosis. In gymnosperms, three of the four haploid spores produced in meiosis typically degenerate, leaving one surviving megaspore inside the nucellus. Among angiosperms, however, a wide range of variation exists in what happens next. The number (and position) of surviving megaspores, the total number of cell divisions, whether nuclear fusions occur, and the final number, position and ploidy of the cells or nuclei all vary. A common pattern of embryo sac development (the Polygonum type maturation pattern) includes a single functional megaspore followed by three rounds of mitosis. In some cases, however, two megaspores survive (for example, in Allium and Endymion). In many cases, all four megaspores survive. For example in Fritillaria type embryo sac development (illustrated by Lilium in the figure) three of the megaspores fuse to form a triploid nucleus. The subsequent arrangement of cells is similar to the Polygonum pattern, but the ploidy of the nuclei is different.
After fertilization, the nucellus may develop into the perisperm that feeds the embryo. In some plants, the diploid tissue of the nucellus can give rise to a seed through a mechanism of asexual reproduction called nucellar embryony.
Megagametophyte
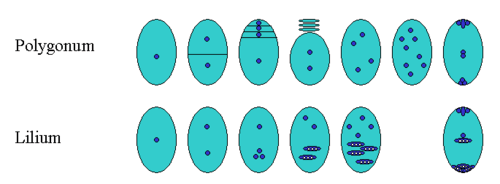

The haploid megaspore inside the nucellus gives rise to the female gametophyte, called the megagametophyte.
In gymnosperms, the megagametophyte consists of around 2000 nuclei and forms archegonia, which produce egg cells for fertilization.
In flowering plants, the megagametophyte (also referred to as the embryo sac) is much smaller and typically consists of only seven cells and eight nuclei. This type of megagametophyte develops from the megaspore through three rounds of mitotic divisions. The cell closest to the micropyle opening of the integuments differentiates into the egg cell, with two synergid cells by its side that are involved in the production of signals that guide the pollen tube. Three antipodal cells form on the opposite (chalazal) end of the ovule and later degenerate. The large central cell of the embryo sac contains two polar nuclei.
Zygote, embryo and endosperm
The pollen tube releases two sperm nuclei into the ovule. In gymnosperms, fertilization occurs within the archegonia produced by the female gametophyte. While it is possible that several egg cells are present and fertilized, typically only one zygote will develop into a mature embryo as the resources within the seed are limited.
In flowering plants, one sperm nucleus fuses with the egg cell to produce a zygote, the other fuses with the two polar nuclei of the central cell to give rise to the polyploid (typically triploid) endosperm. This double fertilization is unique to flowering plants, although in some other groups the second sperm cell does fuse with another cell in the megagametophyte to produce a second embryo. The plant stores nutrients such as starch, proteins, and oils in the endosperm as a food source for the developing embryo and seedling, serving a similar function to the yolk of animal eggs. The endosperm is also called the albumen of the seed.
Types of Embryo Sacs
Embryo sacs may be described according to the number of megaspores developing, as either monosporic, bisporic, or tetrasporic.
See also
References
- ↑ Herr, J.M. Jr., 1995. The orign of the ovule. Am. J. Bot. 82(4):547-64
- ↑ Stewart and Rothwell, 1993. Paleobotany and the evolution of plants. Cambridge University Press.
- ↑ Frohlich and Chase, 2007. After a dozen years of progress, the origin of angiosperms is still a great mystery. Nature 450:1184-1189 (20 December 2007) | doi:10.1038/nature06393;
Further readings
- P.H. Raven, R.F. Evert, S.E. Eichhorn (2005): Biology of Plants, 7th Edition, W.H. Freeman and Company Publishers, New York, ISBN 0-7167-1007-2