Neuroactive steroid
Neuroactive steroids (or neurosteroids) rapidly alter neuronal excitability through interaction with neurotransmitter-gated ion channels.[1][2] In addition, these steroids may also exert effects on gene expression via intracellular steroid hormone receptors. Neurosteroids have a wide range of potential clinical applications from sedation to treatment of epilepsy[3] and traumatic brain injury.[4][5] Ganaxolone, an analog of the endogenous neurosteroid allopregnanolone, is under investigation for the treatment of epilepsy.[6]
Biosynthesis
Several of these steroids accumulate in the brain after local synthesis or after metabolism of adrenal steroids or gonadal steroids, especially testosterone. Neurosteroids are synthesized in the central and peripheral nervous system, especially in myelinating glial cells, from cholesterol or steroidal precursors imported from peripheral sources.[7][8] They include 3β-hydroxy-Δ5 derivatives, such as pregnenolone (PREG) and dehydroepiandrosterone (DHEA), their sulfates, and reduced metabolites such as the tetrahydro derivative of progesterone 3α-hydroxy-5α-pregnane-20-one (3α,5α-THPROG).
Mechanism
These compounds can act as allosteric modulators of neurotransmitter receptors, such as GABAA,[9][10][11][12] NMDA,[13] and sigma receptors.[14] Progesterone (PROG) is also a neurosteroid which activates progesterone receptors expressed in peripheral and central glial cells.[15][16][17][18] The 3α-hydroxy ring A-reduced pregnane steroids allopregnanolone and tetrahydrodeoxycorticosterone have been surmised to enhance GABA-mediated chloride currents, whereas pregnenolone sulfate and dehydroepiandrosterone (DHEA) sulfate display functional antagonistic properties at GABAA receptors.
Therapeutic application
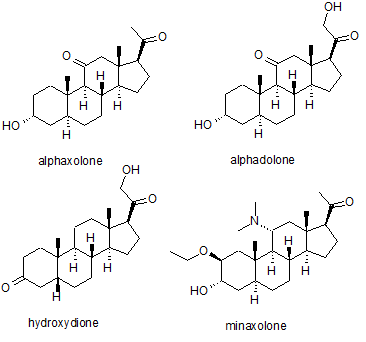
Several synthetic neurosteroids have been used as sedatives for the purpose of general anaesthesia for carrying out surgical procedures. The best known of these are alphaxolone, alphadolone, hydroxydione and minaxolone. The first of these to be introduced was hydroxydione, which is the esterified 21-hydroxy derivative of 5β-pregnanedione. Hydroxydione proved to be a useful anaesthetic drug with a good safety profile, but was painful and irritating when injected probably due to poor water solubility. This led to the development of newer neuroactive steroids. The next drug from this family to be marketed was a mixture of alphaxolone and alphadolone, known as Althesin. This was withdrawn from human use due to rare but serious toxic reactions, but is still used in veterinary medicine. The next neurosteroid anaesthetic introduced into human medicine was the newer drug minaxolone, which is around three times more potent than althesin and retains the favourable safety profile, without the toxicity problems seen with althesin. However this drug was also ultimately withdrawn, not because of problems in clinical use, but because animal studies suggested potential carcinogenicity and since alternative agents were available it was felt that the possible risk outweighed the benefit of keeping the drug on the market.

The neurosteroid ganaxolone, an analog of the progesterone metabolite allopregnanolone, has been extensively investigated in animal models and is currently in clinical trials for the treatment of epilepsy. Neurosteroids, including ganaxolone have a broad spectrum of activity in animal models.[19] They may have advantages over other GABAA receptor modulators, notably benzodiazepines, in that tolerance does not appear to occur with extended use.[20][21] A randomized, placebo controlled, 10 week phase 2 clinical trial of orally administered ganaxolone in adults with partial onset seizure demonstrated that the treatment is safe, well tolerated and efficacious.[6] The drug continued to demonstrate efficacy in an 104 week open label extension. Data from non-clinical studies suggest that ganaxolone may have low risk for use in pregnancy. In addition to use in the treatment of epilepsy, the drug has potential in the treatment of a broad range of neurological and psychiatric conditions. Proof-of-concept studies are currently underway in posttraumatic stress disorder and fragile X syndrome.
Role in antidepressant action
Certain antidepressant drugs such as fluoxetine and fluvoxamine which are generally thought to act primarily as selective serotonin reuptake inhibitors have also been found to increase the levels of certain neurosteroids.[22][23] Based on these studies, it has been proposed that increased levels of neurosteroids induced by fluoxetine or fluvoxamine may significantly contribute to or even be the predominant mechanism of action of these antidepressant drugs.
Benzodiazepine effects on neurosteroids
Benzodiazepines may influence neurosteroid metabolism by virtue of their actions on translocator protein (TSPO; "peripheral benzodiazepine receptor").[24] The pharmacological actions of benzodiazepines at the GABAA receptor are similar to those of neurosteroids. Factors which affect the ability of individual benzodiazepines to alter neurosteroid levels may depend upon whether the individual benzodiazepine drug interacts with TSPO. Some benzodiazepines may also inhibit neurosteroidogenic enzymes reducing neurosteroid synthesis.[25]
Examples
- Allopregnanolone
- Dehydroepiandrosterone
- Dehydroepiandrosterone sulfate
- 5α-Dihydroprogesterone
- Pregnenolone
- Progesterone
- Tetrahydrodeoxycorticosterone
Antagonists
- 17-Phenylandrostenol - blocks the effects of neuroactive steroids without affecting responses produced by benzodiazepines or barbiturates
See also
- 5-alpha reductase
- GABA A receptor
References
- ↑ Paul SM, Purdy RH (1992). "Neuroactive steroids" (abstract). FASEB J. 6 (6): 2311–22. PMID 1347506.
- ↑ Lan NC, Gee KW (1994). "Neuroactive steroid actions at the GABA-A receptor". Horm Behav 28 (4): 537–44. doi:10.1006/hbeh.1994.1052. PMID 7729823.
- ↑ Reddy DS, Rogawski MA (April 2009). "Neurosteroid replacement therapy for catamenial epilepsy". Neurotherapeutics 6 (2): 392–401. doi:10.1016/j.nurt.2009.01.006. PMC 2682439. PMID 19332335.
- ↑ Morrow AL (2007). "Recent Developments in the Significance and Therapeutic Relevance of Neuroactive Steroids – Introduction to the Special Issue". Pharmacol. Ther. 116 (1): 1–6. doi:10.1016/j.pharmthera.2007.04.003. PMC 2047816. PMID 17531324.
- ↑ Dubrovsky BO (2005). "Steroids, neuroactive steroids and neurosteroids in psychopathology". Prog. Neuropsychopharmacol. Biol. Psychiatry 29 (2): 169–92. doi:10.1016/j.pnpbp.2004.11.001. PMID 15694225.
- ↑ 6.0 6.1 Farfel G, Tsai J, Shaw K, Nohria V, Rogawski MA (2013). ""Ganaxolone" in Bialer M, Johannessen SI, Levy RH, Perucca E, Tomson T, White HS: Progress report on new antiepileptic drugs: a summary of the Eleventh Eilat Conference (EILAT XI)". Epilepsy Res. 103 (1): 2–30. doi:10.1016/j.eplepsyres.2012.10.001. PMID 23219031.
- ↑ Agís-Balboa RC, Pinna G, Zhubi A, Maloku E, Veldic M, Costa E, Guidotti A (2006). "Characterization of brain neurons that express enzymes mediating neurosteroid biosynthesis". Proc. Natl. Acad. Sci. U.S.A. 103 (39): 14602–7. doi:10.1073/pnas.0606544103. PMC 1600006. PMID 16984997.
- ↑ Mellon SH, Griffin LD (2002). "Neurosteroids: biochemistry and clinical significance". Trends Endocrinol. Metab. 13 (1): 35–43. doi:10.1016/S1043-2760(01)00503-3. PMID 11750861.
- ↑ Majewska MD, Harrison NL, Schwartz RD, Barker JL, Paul SM (1986). "Steroid hormone metabolites are barbiturate-like modulators of the GABA receptor". Science 232 (4753): 1004–7. doi:10.1126/science.2422758. PMID 2422758.
- ↑ Herd MB, Belelli D, Lambert JJ (2007). "Neurosteroid modulation of synaptic and extrasynaptic GABAA receptors". Pharmacology & Therapeutics 116 (1): 20–34. doi:10.1016/j.pharmthera.2007.03.007. PMID 17531325.
- ↑ Hosie AM, Wilkins ME, da Silva HM, Smart TG (2006). "Endogenous neurosteroids regulate GABAA receptors through two discrete transmembrane sites". Nature 444 (7118): 486–9. doi:10.1038/nature05324. PMID 17108970.
- ↑ Puia G, Santi MR, Vicini S, Pritchett DB, Purdy RH, Paul SM, Seeburg PH, Costa E (1990). "Neurosteroids act on recombinant human GABAA receptors". Neuron 4 (5): 759–65. doi:10.1016/0896-6273(90)90202-Q. PMID 2160838.
- ↑ Wu FS, Gibbs TT, Farb DH (1991). "Pregnenolone sulfate: a positive allosteric modulator at the N-methyl-D-aspartate receptor" (abstract). Mol. Pharmacol. 40 (3): 333–6. PMID 1654510.
- ↑ Maurice T, Junien JL, Privat A (1997). "Dehydroepiandrosterone sulfate attenuates dizocilpine-induced learning impairment in mice via sigma 1-receptors". Behav. Brain Res. 83 (1–2): 159–64. doi:10.1016/S0166-4328(97)86061-5. PMID 9062676.
- ↑ Baulieu EE (1997). "Neurosteroids: of the nervous system, by the nervous system, for the nervous system". Recent Prog. Horm. Res. 52: 1–32. PMID 9238846.
- ↑ Rupprecht R, Reul JM, Trapp T, van Steensel B, Wetzel C, Damm K, Zieglgänsberger W, Holsboer F (1993). "Progesterone receptor-mediated effects of neuroactive steroids". Neuron 11 (3): 523–30. doi:10.1016/0896-6273(93)90156-L. PMID 8398145.
- ↑ Jung-Testas I, Do Thi A, Koenig H, Désarnaud F, Shazand K, Schumacher M, Baulieu EE (1999). "Progesterone as a neurosteroid: synthesis and actions in rat glial cells". J. Steroid Biochem. Mol. Biol. 69 (1–6): 97–107. doi:10.1016/S0960-0760(98)00149-6. PMID 10418983.
- ↑ Belelli D, Lambert JJ (2005). "Neurosteroids: endogenous regulators of the GABAA receptor". Nat. Rev. Neurosci. 6 (7): 565–75. doi:10.1038/nrn1703. PMID 15959466.
- ↑ Rogawski MA, Reddy DS, 2004. Neurosteroids: endogenous modulators of seizure susceptibility. In: Rho, J.M., Sankar, R., Cavazos, J. (Eds.), Epilepsy: Scientific Foundations of Clinical Practice. Marcel Dekker, New York, 2004;319-355.
- ↑ Kokate TG, Yamaguchi S, Pannell LK, et al. (November 1998). "Lack of anticonvulsant tolerance to the neuroactive steroid pregnanolone in mice". J. Pharmacol. Exp. Ther. 287 (2): 553–8. PMID 9808680.
- ↑ Reddy DS, Rogawski MA (December 2000). "Chronic treatment with the neuroactive steroid ganaxolone in the rat induces anticonvulsant tolerance to diazepam but not to itself". J. Pharmacol. Exp. Ther. 295 (3): 1241–8. PMID 11082461.
- ↑ Uzunova V, Sheline Y, Davis JM, Rasmusson A, Uzunov DP, Costa E, Guidotti A (1998). "Increase in the cerebrospinal fluid content of neurosteroids in patients with unipolar major depression who are receiving fluoxetine or fluvoxamine". Proc. Natl. Acad. Sci. U.S.A. 95 (6): 3239–44. doi:10.1073/pnas.95.6.3239. PMC 19726. PMID 9501247.
- ↑ Pinna G, Costa E, Guidotti A (2006). "Fluoxetine and norfluoxetine stereospecifically and selectively increase brain neurosteroid content at doses that are inactive on 5-HT reuptake". Psychopharmacology (Berl.) 186 (3): 362–72. doi:10.1007/s00213-005-0213-2. PMID 16432684.
- ↑ Dhir A, Rogawski MA (October 2011). "Role of neurosteroids in the anticonvulsant activity of midazolam". Br J Pharmacol 165 (8): 2684–91. doi:10.1111/j.1476-5381.2011.01733.x. PMC 3423249. PMID 22014182.
- ↑ Usami N; Yamamoto T, Shintani S, Ishikura S, Higaki Y, Katagiri Y, Hara A (April 2002). "Substrate specificity of human 3(20)alpha-hydroxysteroid dehydrogenase for neurosteroids and its inhibition by benzodiazepines" (pdf). Biol Pharm Bull 25 (4): 441–5. doi:10.1248/bpb.25.441. PMID 11995921.
Further reading
- Akk G, Shu HJ, Wang C, Steinbach JH, Zorumski CF, Covey DF, Mennerick S (2005). "Neurosteroid access to the GABAA receptor". J. Neurosci. 25 (50): 11605–13. doi:10.1523/JNEUROSCI.4173-05.2005. PMID 16354918.
- Wang JM, Johnston PB, Ball BG, Brinton RD (2005). "The neurosteroid allopregnanolone promotes proliferation of rodent and human neural progenitor cells and regulates cell-cycle gene and protein expression". J. Neurosci. 25 (19): 4706–18. doi:10.1523/JNEUROSCI.4520-04.2005. PMID 15888646.
- Dong E, Matsumoto K, Uzunova V, Sugaya I, Takahata H, Nomura H, Watanabe H, Costa E, Guidotti A (2001). "Brain 5α-dihydroprogesterone and allopregnanolone synthesis in a mouse model of protracted social isolation". Proc. Natl. Acad. Sci. U.S.A. 98 (5): 2849–54. doi:10.1073/pnas.051628598. PMC 30228. PMID 11226329.
- Melcangi RC, Celotti F, Martini L (1994). "Progesterone 5-alpha-reduction in neuronal and in different types of glial cell cultures: type 1 and 2 astrocytes and oligodendrocytes". Brain Res. 639 (2): 202–6. doi:10.1016/0006-8993(94)91731-0. PMID 8205473.
- Corpéchot C, Robel P, Axelson M, Sjövall J, Baulieu EE (1981). "Characterization and measurement of dehydroepiandrosterone sulfate in rat brain". Proc. Natl. Acad. Sci. U.S.A. 78 (8): 4704–7. doi:10.1073/pnas.78.8.4704. PMC 320231. PMID 6458035.
- Reddy D, Rogawski MA (2012). "Neurosteroids — Endogenous regulators of seizure susceptibility and role in the treatment of epilepsy". In: Noebels JL, Avoli M, Rogawski MA, et al., editors. Jasper's Basic Mechanisms of the Epilepsies [Internet]. 4th edition. Bethesda (MD): National Center for Biotechnology Information (US); 2012. Available from: http://www.ncbi.nlm.nih.gov/books/NBK98218/. PMID 22787590.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||