Inclusive fitness
| Part of a series on |
| Evolutionary biology |
|---|
 |
|
Natural history |
|
History of evolutionary theory |
|
Fields and applications
|
|
Social implications |
|
In evolutionary biology and evolutionary psychology, the inclusive fitness of an organism is judged by the number of offspring it has, how they support them, and how their offspring could support others.[1] Advocates of inclusive fitness theory say that an organism can improve its overall genetic success by altruistic social behavior.
From the gene's point of view, evolutionary success ultimately depends on leaving behind the maximum number of copies of itself in the population. Until 1964, it was generally believed that genes only achieved this by causing the individual to leave the maximum number of viable offspring. However, in 1964 W. D. Hamilton proved mathematically that, because close relatives of an organism share some identical genes, a gene can also increase its evolutionary success by promoting the reproduction and survival of these related or otherwise similar individuals.
Belding's ground squirrel provides an example. The ground squirrel gives an alarm call to warn its local group of the presence of a predator. By emitting the alarm, it gives its own location away, putting itself in more danger. In the process, however, the squirrel protects its relatives within the local group (along with the rest of the group). Therefore, if protecting the other squirrels in the immediate area will lead to the passing on of more of the squirrel’s own genes than the squirrel could leave by reproducing on its own, then natural selection will favor giving the alarm call, provided that a sufficient fraction of the shared genes include the gene(s) predisposing to the alarm call.[2]
Synalpheus regalis, a eusocial shrimp, also is an example of an organism that seeks to increase its inclusive fitness. The larger defenders protect the young juveniles in the colony from outsiders. By ensuring the young's survival, the genes will continue to be passed on to future generations. [3]
Inclusive fitness is more generalized than strict kin selection, which requires that the shared genes are identical by descent. Inclusive fitness is not limited to cases where kin are involved.
Hamilton's rule
In the context of sociobiology, which holds that some behavior can be influenced by genes and therefore can evolve by natural selection, Hamilton proposed that inclusive fitness offers a mechanism for the evolution of altruism. He claimed that this leads natural selection to favor organisms that would behave in ways that maximize their inclusive fitness.
Hamilton's rule describes mathematically whether or not a gene for altruistic behavior will spread in a population:
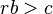
where
-
 is the probability, above the population average, of the individuals sharing an altruistic gene – commonly viewed as "degree of relatedness".
is the probability, above the population average, of the individuals sharing an altruistic gene – commonly viewed as "degree of relatedness". -
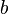 is the reproductive benefit to the recipient of the altruistic behavior, and
is the reproductive benefit to the recipient of the altruistic behavior, and -
 is the reproductive cost to the altruist,
is the reproductive cost to the altruist,
In a recent paper, Gardner et al. suggest that Hamilton's rule can be applied to multi-locus models, but that it should be done at the point of interpreting theory, rather than the starting point of enquiry.[4] They suggest that one should “use standard population genetics, game theory, or other methodologies to derive a condition for when the social trait of interest is favored by selection and then use Hamilton’s rule as an aid for conceptualizing this result". A recent paper by Nowak et al. suggested that standard natural selection theory is superior to inclusive fitness theory, stating that the interactions between cost and benefit can not be explained only in terms of relatedness. This, Nowak said, makes Hamilton's rule at worst superfluous and at best ad hoc.[5] Gardner in turn was critical of the paper, describing it as "a really terrible article", and along with other co-authors has written a reply, submitted to Nature.[6]
In work prior to Nowak, various authors derived different versions of a formula for  , all designed to preserve Hamilton's rule.[7][8][9] Orlove noted that if a formula for is defined so as to ensure that Hamilton's rule is preserved, then the approach is by definition ad hoc. However, he published an unrelated derivation of the same formula for – a derivation designed to preserve two statements about the rate of selection – which on its own was similarly ad hoc. Orlove argued that the existence of two unrelated derivations of the formula for reduces or eliminates the ad hoc nature of the formula, and of inclusive fitness theory as well.[10] The derivations were demonstrated to be unrelated by corresponding parts of the two identical formulae for being derived from the genotypes of different individuals. The parts that were derived from the genotypes of different individuals were terms to the right of the minus sign in the covariances in the two versions of the formula for . By contrast, the terms left of the minus sign in both derivations come from the same source.
, all designed to preserve Hamilton's rule.[7][8][9] Orlove noted that if a formula for is defined so as to ensure that Hamilton's rule is preserved, then the approach is by definition ad hoc. However, he published an unrelated derivation of the same formula for – a derivation designed to preserve two statements about the rate of selection – which on its own was similarly ad hoc. Orlove argued that the existence of two unrelated derivations of the formula for reduces or eliminates the ad hoc nature of the formula, and of inclusive fitness theory as well.[10] The derivations were demonstrated to be unrelated by corresponding parts of the two identical formulae for being derived from the genotypes of different individuals. The parts that were derived from the genotypes of different individuals were terms to the right of the minus sign in the covariances in the two versions of the formula for . By contrast, the terms left of the minus sign in both derivations come from the same source.
Inclusive fitness and altruism
The concept serves to explain how natural selection can perpetuate altruism. If there is an "altruism gene" (or complex of genes) that influences an organism's behavior to be helpful and protective of relatives and their offspring, this behavior also increases the proportion of the altruism gene in the population, because relatives are likely to share genes with the altruist due to common descent. In formal terms, if such a complex of genes arises, Hamilton's rule (rb>c) specifies the selective criteria (in terms of cost, benefit and relatedness) for such a trait to increase in frequency in the population. Hamilton noted that inclusive fitness theory does not by itself predict that a species will necessarily evolve such altruistic behaviors, since an opportunity or context for interaction between individuals is a more primary and necessary requirement in order for any social interaction to occur in the first place. As Hamilton put it, “Altruistic or selfish acts are only possible when a suitable social object is available. In this sense behaviours are conditional from the start.” (Hamilton 1987, 420).[11] In other words, whilst inclusive fitness theory specifies a set of necessary criteria for the evolution of altruistic traits, it does not specify a sufficient condition for their evolution in any given species. More primary necessary criteria include the existence of gene complexes for altruistic traits in gene pool, as mentioned above, and especially that "a suitable social object is available", as Hamilton noted. Paul Sherman, who has contributed much research on the ground squirrels mentioned above, gives a fuller discussion of Hamilton's latter point:
To understand any species’ pattern of nepotism, two questions about individuals’ behavior must be considered: (1) what is reproductively ideal?, and (2) what is socially possible? With his formulation of “inclusive fitness,” Hamilton suggested a mathematical way of answering (1). Here I suggest that the answer to (2) depends on demography, particularly its spatial component, dispersal, and its temporal component, mortality. Only when ecological circumstances affecting demography consistently make it socially possible will nepotism be elaborated according to what is reproductively ideal. For example, if dispersing is advantageous and if it usually separates relatives permanently, as in many birds (Nice 1937: 180-187; Gross 1940; Robertson 1969), on the rare occasions when nestmates or other kin live in proximity, they will not preferentially cooperate. Similarly, nepotism will not be elaborated among relatives that have infrequently coexisted in a population’s or a species’ evolutionary history. If an animal’s life history characteristics (Stearns 1976; Warner this volume) usually preclude the existence of certain relatives, that is if kin are usually unavailable, the rare coexistence of such kin will not occasion preferential treatment. For example, if reproductives generally die soon after zygotes are formed, as in many temperate zone insects, the unusual individual that survives to interact with its offspring is not expected to behave parentally. (Sherman 1980, 530, underlining in original) [12]
The occurrence of sibling cannibalism in several species[13][14][15] underlines the point that inclusive fitness theory should not be understood to simply predict that genetically related individuals will inevitably recognize and engage in positive social behaviors towards genetic relatives. Only in species that have the appropriate traits in their gene pool, and in which individuals typically interacted with genetic relatives in the natural conditions of their evolutionary history, will social behavior potentially be elaborated, and consideration of the evolutionarily typical demographic composition of grouping contexts of that species is thus a first step in understanding how selection pressures upon inclusive fitness have shaped the forms of its social behavior. Dawkins gives a simplified illustration:
If families [genetic relatives] happen to go around in groups, this fact provides a useful rule of thumb for kin selection: ‘care for any individual you often see’.” (Dawkins 1979, 187)[16]
Evidence from a variety of species [17][18][19] including primates[20] and other social mammals[21] suggests that contextual cues (such as familiarity) are often significant proximate mechanisms mediating the expression of altruistic behavior, regardless of whether the participants are always in fact genetic relatives or not. This is nevertheless evolutionarily stable since selection pressure acts on the typical conditions, not on the rare occasions where actual genetic relatedness differs from that normally encountered (see Sherman above). Inclusive fitness theory thus does not imply that organisms evolve to direct altruism towards genetic relatives. Many popular treatments do however promote this interpretation, as illustrated in a recent review:
[M]any misunderstandings persist. In many cases, they result from conflating “coefficient of relatedness” and “proportion of shared genes,” which is a short step from the intuitively appealing—but incorrect—interpretation that “animals tend to be altruistic toward those with whom they share a lot of genes.” These misunderstandings don’t just crop up occasionally; they are repeated in many writings, including undergraduate psychology textbooks—most of them in the field of social psychology, within sections describing evolutionary approaches to altruism. (Park 2007, p860)[22]
Such misunderstandings of inclusive fitness' implications for the study of altruism, even amongst professional biologists utilizing the theory, are widespread, prompting prominent theorists to regularly attempt to highlight and clarify the mistakes.[16] Here is one recent example of attempted clarification from West et al. (2010):
In his original papers on inclusive fitness theory, Hamilton pointed out a sufficiently high relatedness to favour altruistic behaviours could accrue in two ways —kin discrimination or limited dispersal ( Hamilton, 1964, 1971,1972, 1975). There is a huge theoretical literature on the possible role of limited dispersal reviewed by Platt & Bever (2009) and West et al. (2002a), as well as experimental evolution tests of these models (Diggle et al., 2007; Griffin et al., 2004; Kümmerli et al., 2009 ). However, despite this, it is still sometimes claimed that kin selection requires kin discrimination (Oates & Wilson, 2001; Silk, 2002 ). Furthermore, a large number of authors appear to have implicitly or explicitly assumed that kin discrimination is the only mechanism by which altruistic behaviours can be directed towards relatives... [T]here is a huge industry of papers reinventing limited dispersal as an explanation for cooperation. The mistakes in these areas seem to stem from the incorrect assumption that kin selection or indirect fitness benefits require kin discrimination (misconception 5), despite the fact that Hamilton pointed out the potential role of limited dispersal in his earliest papers on inclusive fitness theory (Hamilton, 1964; Hamilton, 1971; Hamilton, 1972; Hamilton, 1975). (West et al. 2010, p.243 and supplement)[23]
Green-beard effects
As well as interactions within reliable contexts of genetic relatedness, altruists may also have some way to recognize altruistic behavior in unrelated individuals and be inclined to support them. As Dawkins points out in The Selfish Gene (Chapter 6) and The Extended Phenotype,[24] this must be distinguished from the green-beard effect. The proposal that altruists who support other altruists who are not their kin will encourage the evolution of altruism requires that altruists recognize and choose to support others predisposed toward altruism whom they have detected by their past altruistic behavior, not on the observation of some temporarily correlated characteristic (e.g., altruists have green beards). If the green beard effect were the mechanism, some non-altruistic individuals would evolve to mimic the label and would receive the benefits of support from altruists. This would happen quickly due to crossing over of chromosomes; it would not require waiting for the rare event of a mutation. The mimics would receive the benefits but would not incur the costs of caring for others, and so would out-compete the true altruists. Green-Beard transfers thus have a negative affect on the evolution of altruistic behaviors.
Inclusive fitness and parental care
Some might express concern that parental investment (parental care) is said to contribute to inclusive fitness. The distinctions between the kind of beneficiaries nurtured (collateral versus descendant relatives) and the kind of fitnesses used (inclusive versus personal) in the parsing of nature are orthogonal concepts. This orthogonality can best be understood in a thought experiment: Consider a model of a population of animals such as crocodiles or tangle web spiders. Some species or populations of these spiders and reptiles exhibit parental care, while closely related species or populations lack it. Assume that in these animals a gene, called a, codes for parental care, and its other allele, called A, codes for an absence thereof. The aa homozygotes care for their young, and AA homozygotes don't, and the heterozygotes behave like aa homozygotes if a is dominant, and like AA homozygotes if A is dominant, or exhibit some kind of intermediate behavior if there is partial dominance. Other kinds of animals could be considered in which all individuals exhibit parental care, but variation among them would be in the quantity and quality thereof.
If one considers a life cycle as extending from conception to conception, and an animal is an offspring of parents with poor parental care, then the higher mortality with poor care could be considered a diminution of the offspring's expected fitness.
Alternatively, if one considers the life cycle as extending from weaning to weaning, the same mortality would be considered a diminution in the parents' fecundity, and therefore a diminution of the parent's fitness.
In Hamilton's paradigm, fitnesses calculated according to in the weaning-to-weaning perspective are inclusive fitnesses, and fitnesses calculated in the conception-to-conception perspective are personal fitnesses. This distinction is independent of whether the altruism involved in child rearing is toward descendents or toward collateral relatives, as when aunts and uncle rear their nieces and nephews.
Inclusive fitness theory was developed in order to better understand collateral altruism, but this does not mean that it is limited to collateral altruism. It applies just as well to parental care. Which perspective one chooses does not affect the animals but just one's understanding.
See also
- Gene-centered view of evolution
- Evolutionary psychology
- Criticism of evolutionary psychology
- Kin selection
- Reproductive success
- r/K selection theory
References
- ↑ Definition and explanation of inclusive fitness from Personality Research.org
- ↑ Mateo JM, 1996. The development of alarm-call response behavior in free-living juvenile Belding's ground squirrels. Animal Behaviour 52:489-505.
- ↑ J. Emmett Duffy, Cheryl L. Morrison & Kenneth S. Macdonald (2002). "Colony defense and behavioral differentiation in the eusocial shrimp Synalpheus regalis" (PDF). Behavioral Ecology and Sociobiology 51 (5): 488–495. doi:10.1007/s00265-002-0455-5.
- ↑ Gardner A, West SA and Barton NH. The relation between multilocus population genetics and social evolution theory. Am Nat 169, 207–226.
- ↑ Martin A. Nowak, Corina E. Tarnita & Edward O. Wilson The evolution of eusociality Nature 466 1057–1062
- ↑ http://www.nytimes.com/2010/08/31/science/31social.html?pagewanted=1&_r=1
- ↑ Orlove, M. J. 1975 A Model of Kin Selection not Invoking Coefficients of Relationship J. Theor. Biol. v49 pp289-310
- ↑ Orlove, M. J. & Wood, C. L. 1978. "Coefficients of relationship and coefficients of relatedness in kin selection: A covariance form for the RHO formula". Journal of Theoretical Biology, Volume 73, Issue 4, 21 August 1978, Pages 679-686
- ↑ Michod, R. E. & Hamilton, W. D. 1980. "Coefficients of relatedness in sociobiology" Nature 288, 694 - 697 (18 December 1980)
- ↑ Orlove, M. J. 1979 A Reconciliation of Inclusive Fitness and Personal Fitness Approaches: a Proposed Correcting Term for the Inclusive Fitness Formula, J. Theor. Biol. v81 pp 577–586
- ↑ Hamilton, W.D. 1987. Discriminating nepotism: expectable, common and overlooked. In Kin recognition in animals, edited by D. J. C. Fletcher and C. D. Michener. New York: Wiley.
- ↑ Sherman, P. W. 1980. The limits of ground squirrel nepotism. In Sociobiology: beyond nature/nurture?, edited by G. W. Barlow and J. Silverberg. Boulder, Colorado: Westview Press.
- ↑ Summers, K., and R. Symula. 2001. Cannibalism and kin discrimination in tadpoles of the amazonian poison frog, Dendrobates ventrimaculatus, in the field. Herpetological Journal 11 (1):17–21.
- ↑ Gabor, C. R. 1996. Differential kin discrimination by red-spotted newts (Notophthalmus viridescens) and smooth newts (Triturus vulgaris). Ethology 102 (8):649-659.
- ↑ Walls, S. C., and A. R. Blaustein. 1995. Larval Marbled Salamanders, Ambystoma-Opacum, Eat Their Kin. Animal Behaviour 50:537-545.
- ↑ 16.0 16.1 Dawkins, R. 1979. 12 Misunderstandings of kin selection. Z. Tierpsychology 51:184-200.
- ↑ Tai, F. D., T. Z. Wang, and Y. J. Zhao. 2000. Inbreeding avoidance and mate choice in the mandarin vole (Microtus mandarinus). Canadian Journal of Zoology-Revue Canadienne De Zoologie 78(12):2119-2125.
- ↑ Hare, J. F., and J. O. Murie. 1996. Ground squirrel sociality and the quest for the 'holy grail': Does kinship influence behavioral discrimination by juvenile Columbian ground squirrels. Behavioral Ecology 7 (1):76-81.
- ↑ Waldman, B. 1988. The Ecology of Kin Recognition. Annual Review of Ecology and Systematics 19:543-571.
- ↑ Silk, J. B. 2001. Ties that Bond: The Role of Kinship in Primate Societies. In New Directions in Anthropological Kinship, edited by L. Stone. Oxford: Rowman and Littlefield.
- ↑ Stookey, J. M., and H. W. Gonyou. 1998. Recognition in swine: recognition through familiarity or genetic relatedness? Applied Animal Behaviour Science 55 (3-4):291-305.
- ↑ Park, J. H. 2007. Persistent Misunderstandings of Inclusive Fitness and Kin Selection: Their Ubiquitous Appearance in Social Psychology Textbooks. Evolutionary Psychology 5(4): 860-873
- ↑ West et al. 2011. Sixteen common misconceptions about the evolution of cooperation in humans. Evolution and Social Behaviour 32 (2011) 231-262
- ↑ Dawkins, Richard, "The Extended Phenotype", Oxford University Press 1982 (Chapter 9)
Sources
- Campbell, N., Reece, J., et al. 2002. Biology. 6th ed. San Francisco, California. pp. 1145–1148.
- Rheingold, Howard, “Technologies of cooperation” in Smart Mobs. Cambridge, MA : Perseus Publishing, 2002 (Ch. 2:pp 29–61)
- Dawkins, Richard C. 1976 The Selfish Gene, Oxford University Press (Discussion of carers and bearers in relation to inclusive and personal fitnesses, and the bugbear of parental investment as part of inclusive fitness occurs herein)
- Hamilton, W. D. 1964 The Genetical Evolution of Social Behaviour I and II, J. Theor. Biol. v7, pp 1–16, and 17-52
- Hamilton, W. D. 1975, Innate Social Aptitudes of Man: an Approach from Evolutionary Genetics, in Robin Fox (ed.), Biosocial Anthropology, Malaby Press, London, 133-153 (IF including altruism to fellow altruists among strangers discussed herein)
- Hamilton, W. D. Narrow Roads of Geneland I and II, 1995 Freeman I 2001 Oxford Press II (biography of WDH and anthology of his writings)
- Orlove, M. J. 1975 A Model of Kin Selection not Invoking Coefficients of Relationship J. Theor. Biol. v49 pp289–310 (Isomorphism between Karma and Kin Theories discussed herein)
- Orlove, M. J. 1979 A Reconciliation of Inclusive Fitness and Personal Fitness Approaches: a Proposed Correcting Term for the Inclusive Fitness Formula, J. Theor. Biol. v81 pp577–586 (Karma-Theory/Kin-Theory equivalence moves from conjecture to theorem status here)
- Trivers, R. L. 1971 The Evolution of Reciprocal Altruism, Quarterly Review of Biology 46: 35-57
- Trivers, R. L. 1972 Parental Investment and Sexual Selection in B. Campbell (ed.), Sexual Selection and the Descent of Man, 1871-1971 (pp. 136–179) Chicago, Il: Aldine
- Trivers, R. L. 1974 Parent/Offspring Conflict, American Zoologist, 14 249-264 (Bigtime importance of If in understanding intra-family conflict)
- Sherman, P.W. 2001. “Squirrels” (pp. 598–609, with L. Wauters) and “The Role of Kinship” (pp. 610–611) in Encyclopedia of Mammals, D.W. Macdonald (Ed.). Andromeda, UK.
External links
- Test simulations: