Dopamine
| Dopamine | |
|---|---|
 | |
 | |
| IUPAC name 4-(2-aminoethyl)benzene-1,2-diol | |
| Other names 2-(3,4-dihydroxyphenyl)ethylamine; | |
| Identifiers | |
| CAS number | 51-61-6 |
| PubChem | 681 |
| ChemSpider | 661 |
| UNII | VTD58H1Z2X |
| DrugBank | DB00988 |
| KEGG | D07870 |
| ChEBI | CHEBI:18243 |
| ChEMBL | CHEMBL59 |
| ATC code | C01 |
| Jmol-3D images | Image 1 |
| |
| |
| Properties | |
| Molecular formula | C8H11NO2 |
| Molar mass | 153.18 g/mol |
| Appearance | colorless solid |
| Density | 1.26 g/cm3 |
| Melting point | 128 °C; 262 °F; 401 K |
| Boiling point | decomposes |
| Solubility in water | 60.0 g/100 ml |
| Except where noted otherwise, data are given for materials in their standard state (at 25 °C (77 °F), 100 kPa) | |
| Infobox references | |
Dopamine (or 3,4-dihydroxyphenethylamine) is a neurotransmitter in the catecholamine and phenethylamine families that plays a number of important roles in the brains and bodies of animals. Its name derives from its chemical structure: it is an amine that is formed by removing a carboxyl group from a molecule of L-DOPA.
In the brain, dopamine functions as a neurotransmitter—a chemical released by nerve cells to send signals to other nerve cells. The brain includes several distinct dopamine systems, one of which plays a major role in reward-motivated behavior. Every type of reward that has been studied increases the level of dopamine in the brain, and a variety of addictive drugs, including stimulants such as cocaine, amphetamine, and methamphetamine, act by amplifying the effects of dopamine. Other brain dopamine systems are involved in motor control and in controlling the release of several important hormones.
Several important diseases of the nervous system are associated with dysfunctions of the dopamine system. Parkinson's disease, a degenerative condition causing tremor and motor impairment, is caused by loss of dopamine-secreting neurons in the midbrain area called the substantia nigra. There is evidence that schizophrenia involves altered levels of dopamine activity, and the antipsychotic drugs that are frequently used to treat it have a primary effect of attenuating dopamine activity. Attention deficit hyperactivity disorder (ADHD) and restless legs syndrome (RLS) are also believed to be associated with decreased dopamine activity.
Outside the nervous system, dopamine functions in several parts of the body as a local chemical messenger. In the blood vessels, it inhibits norepinephrine release[citation needed] and acts as a vasodilator; in the kidneys, it increases sodium excretion and urine output; in the pancreas, it reduces insulin production; in the digestive system, it reduces gastrointestinal motility and protects intestinal mucosa; and in the immune system, it reduces the activity of lymphocytes. With the exception of the blood vessels, dopamine in each of these peripheral systems has a "paracrine" function: it is synthesized locally and exerts its effects on cells that are located near the cells that release it.
A variety of important drugs work by altering the way the body makes or uses dopamine. Dopamine itself is available for intravenous injection: although it cannot reach the brain from the bloodstream, its peripheral effects make it useful in the treatment of heart failure or shock, especially in newborn babies. L-DOPA, the metabolic precursor of dopamine, does reach the brain and is the most widely used treatment for Parkinson's disease. Dopamine-activating stimulants such as cocaine, amphetamine, and methylphenidate (Ritalin)[citation needed] are addictive in high doses, but are used at lower doses to treat ADHD. Conversely, many antipsychotic drugs act by suppressing the effects of dopamine. Drugs that act against dopamine by a different mechanism are also some of the most effective anti-nausea agents.
Dopaminergic systems of the body
In the brain

Inside the brain, dopamine plays important roles in motor control, motivation, arousal, cognition, and reward, as well as a number of basic lower-level functions including lactation, sexual gratification, and nausea.
Dopaminergic neurons (i.e., neurons whose primary neurotransmitter is dopamine) are comparatively few in number — a total of around 400,000 in the human brain[1] — and their cell bodies are confined to a few relatively small brain areas, but they send projections to many other brain areas and exert powerful effects on their targets. These dopaminergic cell groups were first mapped in 1964 by Annica Dahlström and Kjell Fuxe, who assigned them labels starting with the letter "A" (for "aminergic").[2] In their scheme, areas A1 through A7 contain the neurotransmitter norepinephrine, whereas A8 through A14 contain dopamine. Here is a list of the dopaminergic areas they identified:
- The substantia nigra, a small midbrain area that forms a component of the basal ganglia. The dopamine neurons are found mainly in a part of this structure called the pars compacta (cell group A8) and nearby (group A9).[3] In rodents, their most important projections go to the striatum, globus pallidus, and subthalamic nucleus, all of which also belong to the basal ganglia, and play important roles in motor control. The name substantia nigra is Latin for "dark substance", and refers to the fact that the dopaminergic neurons there are darkly pigmented. These neurons are especially vulnerable to damage. When a large fraction of them die, the result is a Parkinsonian syndrome.[4]
- The ventral tegmental area (VTA), another midbrain area. This cell group (A10) is the largest group of dopaminergic cells in the human brain, though still quite small in absolute terms. Projections from these dopaminergic neurons go to the nucleus accumbens and the prefrontal cortex as well as several other areas.[3] These neurons play a central role in reward and other aspects of motivation. The nucleus accumbens is often considered to be the "limbic" part of the striatum. As such, it is the part of the striatum involved in the highest level aspects of motor control, which include motivation and decision-making. Thus, the role of the VTA in motivation and decision-making is structurally analogous to the role of the substantia nigra in low-level motor control.[5] In primates (i.e. monkeys and humans), the dopamine neurons from the regions of the substantia nigra and VTA project throughout most of the cortical mantle, with particularly dense innervation of the motor and premotor cortices. Thus, there are major species differences in cortical dopamine projections.[6]
- The posterior hypothalamus. These dopaminergic cells (group A11) project to the spinal cord, and their function is not well established. There is some evidence that pathology in this area plays a role in restless legs syndrome, a condition in which people have difficulty sleeping due to an overwhelming compulsion to constantly move parts of the body, especially the legs.[7]
- The arcuate nucleus (cell group A12) and periventricular nucleus (cell group A14) of the hypothalamus. An important projection from these dopaminergic neurons goes to the pituitary gland, where it influences the secretion of the hormone prolactin. Dopamine is the primary neuroendocrine inhibitor of the secretion of prolactin from the anterior pituitary gland. Dopamine produced by neurons in the arcuate nucleus is secreted into the hypothalamo-hypophysial blood vessels of the median eminence, which supply the pituitary gland. The lactotrope cells that produce prolactin, in the absence of dopamine, secrete prolactin continuously; dopamine inhibits this secretion. Thus, in the context of regulating prolactin secretion, dopamine is occasionally called prolactin-inhibiting factor (PIF), prolactin-inhibiting hormone (PIH), or prolactostatin.[8]
- The zona incerta. These cells (group A13) project to several areas of the hypothalamus, and participate in the control of gonadotropin-releasing hormone, which is necessary to activate the development of reproductive systems that occurs following puberty, both in males and females.[8]
An additional group of dopamine-secreting neurons are located in the retina of the eye. These neurons are amacrine cells, meaning that they have no axons. They release dopamine into the extracellular medium, and are specifically active during daylight hours, becoming silent at night. This retinal dopamine acts to enhance the activity of cone cells in the retina while suppressing rod cells — the result is to increase sensitivity to color and contrast during bright light conditions, at the cost of reduced sensitivity when the light is dim.[9]
Outside the nervous system
Dopamine does not cross the blood–brain barrier, so its synthesis and functions in peripheral areas are to a large degree independent of its synthesis and functions in the brain. A substantial amount of dopamine circulates in the bloodstream, but its functions there are not entirely clear. Dopamine is found in blood plasma at levels comparable to those of epinephrine, but in humans, over 95% of the dopamine in the plasma is in the form of dopamine sulphate, a conjugate produced by the enzyme Sulfotransferase 1A3/1A4 acting on free dopamine. The bulk of this dopamine sulphate is produced in the mesentric organs that surround parts of the digestive system. The production of dopamine sulphate is thought to be a mechanism for detoxifying dopamine that is ingested as food or produced by the digestive process — plasma levels typically rise more than fifty-fold after a meal. Dopamine sulphate has no known biological functions and is excreted in urine.[10]
The relatively small quantity of unconjugated dopamine in the bloodstream may be produced by the sympathetic nervous system, the digestive system, or possibly other organs. It may act on dopamine receptors in peripheral tissues, or be metabolized, or be converted to norepinephrine by the enzyme dopamine beta hydroxylase, which is released into the bloodstream by the adrenal medulla.[10] Some dopamine receptors are located in the walls of arteries, where they act as a vasodilator and an inhibitor of norepinephrine release.[11] These responses might be activated by dopamine released from the carotid body under conditions of low oxygen, but whether arterial dopamine receptors perform other biologically useful functions is not known.
Beyond its role in modulating blood flow, there are several peripheral systems in which dopamine circulates within a limited area and performs an exocrine or paracrine function.[10] The peripheral systems in which dopamine plays an important role include:
- The immune system. Dopamine acts upon receptors present on immune cells, especially lymphocytes.[12] Dopamine can also affect immune cells in the spleen, bone marrow, and circulatory system.[13] In addition, dopamine can be synthesized and released by immune cells themselves.[12] The main effect of dopamine on lymphocytes is to reduce their activation level. The functional significance of this system is unclear, but it afford a possible route for interactions between the nervous system and immune system, and may be relevant to some autoimmune disorders.[14]
- The kidneys. Multiple types of dopamine receptors are present in cells of the kidneys. Dopamine is also synthesized there, by tubule cells, and discharged into the tubular fluid. Its actions include increasing the blood supply to the kidneys, increasing filtration by the glomeruli, and increasing excretion of sodium in the urine. Defects in renal dopamine function can be produced by high blood pressure or by genetic problems, and can lead to reduced sodium excretion as well as hypertension.[15]
- The pancreas. The role of dopamine here is somewhat complex. The pancreas consists of two parts, known as exocrine and endocrine. The exocrine part synthesizes enzymes and other substances, and secretes them into the small intestine, where food is digested. One of the substances synthesized and secreted by the exocrine pancreas is dopamine. The function of this secreted dopamine after it enters the small intestine is not clearly established — the possibilities include protecting the intestinal mucosa from damage and reducing gastrointestinal motility (the rate at which food moves through the intestines).[16]
- The endocrine part of the pancreas, also known as the islets of Langerhans, synthesizes a number of hormones, including insulin, and secretes them into the bloodstream. There is evidence that the beta cells that synthesize insulin contain dopamine receptors, and that dopamine acts to reduce the amount of insulin they release. The source of their dopamine input is not clearly established — it may come from dopamine that circulates in the bloodstream and derives from the sympathetic nervous system, or it may be synthesized locally by other types of pancreatic cells.[16]
Cellular effects
| Family | Receptor | Gene | Type | Mechanism |
|---|---|---|---|---|
| D1-like | D1 | DRD1 | Gs-coupled. | Increase intracellular levels of cAMP by activating adenylate cyclase. |
| D5 | DRD5 | |||
| D2-like | D2 | DRD2 | Gi/Go-coupled. | Decrease intracellular levels of cAMP by inhibiting adenylate cyclase. |
| D3 | DRD3 | |||
| D4 | DRD4 |
Like many other biologically active substances, dopamine exerts its effects by binding to and activating receptors located on the surface of cells. In mammals, five subtypes of dopamine receptors have been identified, labeled D1 through D5. All of them function as G protein-coupled receptors, meaning that they exert their effects via a complex second messenger system. Glossing over the details, dopamine receptors in mammals can be divided into two families, known as D1-like and D2-like. The ultimate effect of D1-like receptors (D1 and D5) can be excitation (via opening of sodium channels) or inhibition (via opening of potassium channels); the ultimate effect of D2-like receptors (D2, D3, and D4) is usually inhibition of the target neuron. Consequently, it is incorrect to describe dopamine itself as either excitatory or inhibitory. Its effect on a target neuron depends on which types of receptors are present on the membrane of that neuron and on the internal responses of that neuron to cyclic AMP. D1 receptors are the most numerous dopamine receptors in the central nervous system; D2 receptors are next; D3, D4, and D5 receptors are present at significantly lower levels.
The level of extracellular dopamine is modulated by two mechanisms: tonic and phasic dopamine transmission. Tonic dopamine transmission occurs when small amounts of dopamine are released independently of neuronal activity, and is regulated by the activity of other neurons and neurotransmitter reuptake.[17] Phasic dopamine release results from the activity of the dopamine-containing cells themselves. This activity is characterized by irregular pacemaking activity of single spikes, and rapid bursts of typically 2-6 spikes in quick succession.[18][19]
The substantia nigra dopamine system and motor control

The substantia nigra is a component of the basal ganglia, a group of interconnected structures in the forebrain and midbrain that play a central role in motor control. The precise nature of that role has been difficult to work out, but one popular line of thought describes it as "response selection". The response selection theory proposes that when a person or animal is in a situation where several behaviors are possible, activity in the basal ganglia determines which of them is executed, by releasing that response from inhibition. Thus the basal ganglia are responsible for initiating behaviors but not for determining the details of how they are carried out.
Dopamine is thought to modulate the response selection process in at least two important ways. First, dopamine sets the "effort threshold" for initiating behaviors. The higher the level of dopamine activity, the lower the impetus required to evoke a given behavior. As a consequence, high levels of dopamine lead to high levels of motor activity and "impulsive" behavior; low levels of dopamine lead to torpor and slowed reactions. Parkinson's disease, in which dopamine levels in the substantia nigra circuit are greatly reduced, is characterized by stiffness and greatly reduced movement—however, when people with the disease are confronted with strong stimuli such as a serious threat, their reactions can be as vigorous as those of a healthy person. In the opposite direction, drugs that increase the effects of dopamine, such as cocaine or amphetamine, produce heightened levels of activity, including at the highest levels psychomotor agitation and stereotyped movements.
The second important effect of dopamine is as a "teaching" signal. When a motor response is followed by an increase in dopamine activity, the basal ganglia circuit is altered in a way that makes the same response easier to evoke when similar situations arise in the future. This is a form of operant conditioning, in which dopamine plays the role of a reward signal.
Anatomy and physiology
The anatomy of the basal ganglia is extraordinarily complex, and the role of dopamine there is correspondingly complex. On a macroscopic scale there is only one major dopamine projection, from the substantia nigra pars compacta to the striatum, but the dopamine inputs contact multiple types of neurons and have several distinct effects on their targets, activating some via D1 receptors while inhibiting others via D2 receptors. A substantial number of dopamine inputs are delivered to the necks of dendritic spines, where they are well-placed to exert a gating effect on specific synaptic connections, often arising from the cerebral cortex. There are two distinct pathways of signal flow arising from the striatum, known as the direct pathway and indirect pathway. Dopamine is thought to promote action by upregulating the direct pathway while suppressing the indirect pathway.
Many theoreticians believe that the mechanism underlying motor learning in the basal ganglia involves a form of long-term potentiation that occurs in the striatum and is strongly modulated by dopamine—in other words, a mechanism by which dopamine activity induces strengthening or weakening of synaptic connections inside the striatum.[20]
The ventral tegmental area, reward, and cognition
The ventral tegmental area (VTA) contains the largest group of dopamine neurons in the human brain. They project to numerous brain areas, but the two largest projections are the mesolimbic pathway, which targets the nucleus accumbens and other limbic structures, and the mesocortical pathway, which targets the prefrontal and insular parts of the cerebral cortex.
Reward
The VTA dopamine system is strongly associated with the reward system of the brain. Dopamine is released in areas such as the nucleus accumbens and prefrontal cortex as a result of rewarding experiences such as food, sex, and neutral stimuli that become associated with them.[21] The source of this dopamine is primarily the VTA, although the substantia nigra may also contribute. Electrical stimulation of the VTA or its output pathways can itself serve as a potent reward: animals will quickly learn to press a lever if it results in stimulation of dopamine release, and often will continue pressing the lever for a long time, at steadily increasing rates.[22] A variety of drugs that increase dopamine levels are intrinsically rewarding and increase the effects of other types of reward.[22]
In spite of the overwhelming evidence showing a strong association between dopamine and reward, there has been a great deal of dispute about whether the function of dopamine should be described as reward per se, or as some more complex construct that relates strongly to reward. The difficulty arises mainly from two observations: (1) in addition to being rewarding, dopamine is also arousing — it produces a general increase in movement of all sorts; (2) dopamine release can be caused by events that do not seem to have anything to do with reward, most notably pain. One of the most popular alternatives to the reward theory is the "incentive salience" theory, which argues that the function of dopamine is to increase the effects of motivators of all sorts, both positive and negative.[23]
A substantial body of evidence suggests that dopamine encodes not reward itself, but rather reward prediction error, that is, the degree to which reward is surprising. According to this hypothesis, which derives initially from recordings made by Wolfram Schultz, rewards that are expected do not produce any activation of dopamine cells, but rewards that are greater than expected produce a short-lasting increase in dopamine, whereas the omission of an expected reward actually causes dopamine release to drop below its ordinary background level. The "prediction error" hypothesis has drawn particular interest from computational neuroscientists, because an influential computational-learning method known as temporal difference learning makes heavy use of a signal that encodes prediction error. This confluence of theory and data has led to a fertile interaction between theoretical and empirical neuroscientists.[23]
Recent research finds that while some dopaminergic neurons react in the way expected of reward neurons, others do not and seem to respond in regard to salience, including aversive stimuli.[24] This research finds the reward neurons predominate in the ventromedial region of the substantia nigra pars compacta, as well as in the ventral tegmental area. Neurons in these areas project mainly to the ventral striatum and thus might transmit value-related information in regard to reward values.[24] The salience neurons are predominate in the dorsolateral area of the substantia nigra pars compacta which projects to the dorsal striatum and may relate to orienting behaviour.[24] It has been suggested that the difference between these two types of dopaminergic neurons arises from their input: reward-linked ones have input from the basal forebrain, while the salience-related ones from the lateral habenula.[24] In primates, neurons from the regions of both the substantia nigra and VTA project to the prefrontal cortex;[25] the origins of the dopamine innervation of other cortical areas in primate have not been studied. It has been appreciated for many years that exposure to even mild, uncontrollable stress increases dopamine release in the rodent prefrontal cortex, e.g. reviewed in,[26] suggesting that dopamine salience cells have a large influence on this cortical region.
"Seeking" versus "liking"
Kent Berridge and other researchers have argued for a distinction between reward, which is defined in terms of motivation, and pleasure, which is defined in terms of emotional expression. A simpler way of describing this is as a distinction between "seeking" and "liking". "Seeking" occurs when an animal, given access to some stimulus such as food, executes some type of active behavior in order to acquire it. "Liking" occurs when an animal shows expressions of happiness or satisfaction while consuming something. There is considerable evidence that the dopamine system is part of the brain system that mediates seeking but not part of the system that mediates liking. Drugs that increase the effects of dopamine (most notably stimulants such as methamphetamine or cocaine) produce corresponding increases in seeking behaviors, but do not greatly alter expressions of pleasure. Conversely, opiate drugs such as heroin or morphine produce increases in expressions of pleasure but do not greatly alter seeking behaviors. Animals in which the VTA dopamine system has been rendered inactive do not seek food, and will starve to death if left to themselves, but if food is placed in their mouths they will consume it and show facial expressions indicative of pleasure.[21]
Role in cognition
Dopamine's effects on higher cognitive function have been studied in monkeys and rodents. This work began with the landmark study of Brozoski et al., 1979 showing that depletion of catecholamines from the dorsolateral prefrontal cortex in monkeys impaired spatial working memory to the same degree as removing the cortex itself.[27] It is now known that both dopamine and norepinephrine have essential actions on prefrontal cortical function, and help coordinate cognitive state with arousal state.[28] Dopamine has an "inverted U" influence on prefrontal function through its actions on D1 receptors, where either too little or too much impairs working memory function.[29] In the primate prefrontal cortex, dopamine D1 receptor stimulation selectively influences the firing of "Delay" cells (also called "Memory" cells), while dopamine D2 receptors selectively alter the firing of "Response cells".[30]
Diseases and disorders
The dopamine system plays a central role in a number of important medical conditions, including Parkinson's disease, attention deficit hyperactivity disorder, schizophrenia, and drug addiction.
Parkinson's disease
Parkinson's disease is a disorder characterized by stiffness of the body, slowing of movement, and trembling of limbs when they are not in use. In advanced stages it progresses to dementia and eventually death. The main symptoms are caused by massive loss of dopamine-secreting cells in the substantia nigra. These dopamine cells are especially vulnerable to damage, and a variety of insults, including encephalitis (as depicted in the book and movie "Awakenings"), repeated sports-related concussions, and some forms of chemical poisoning (ex. MPTP), can lead to substantial cell loss, producing a Parkinsonian syndrome that is similar in its main features to Parkinson's disease. Most cases of Parkinson's disease, however, are "idiopathic", meaning that the cause of cell death cannot be identified.
The most widely used treatment for Parkinsonism is administration of L-DOPA, the metabolic precursor for dopamine. This treatment cannot restore the dopamine cells that have been lost, but it causes the remaining cells to produce more dopamine, thereby compensating for the loss to at least some degree. In advanced stages the treatment begins to fail because the cell loss is so severe that the remaining ones cannot produce enough dopamine regardless of L-DOPA levels. As this stage is approached, the metabolic regulatory mechanisms in the dopamine cells, operating far above their normal level, become erratic, producing dopamine dysregulation syndrome, in which patients fluctuate unpredictably between states of hyperactivity and paralysis.[31]
Attention deficit hyperactivity disorder
Altered dopamine neurotransmission is implicated in attention deficit hyperactivity disorder (ADHD), a condition associated with impaired ability to regulate attention, behavior, and/or impulses. There are some genetic links between dopamine receptors, the dopamine transporter and ADHD,[32] in addition to links to other neurotransmitter receptors and transporters. The most important relationship between dopamine and ADHD involves the drugs that are used to treat ADHD. Some of the most effective therapeutic agents for ADHD are psychostimulants such as methylphenidate (Ritalin) and amphetamine, drugs that increase both dopamine and norepinephrine levels in brain.[33]
Drug addiction
A variety of addictive drugs produce an increase in reward-related dopamine activity. For some addictive drugs such as alcohol or heroin, activation of the reward system may play only a minor role in addiction, with suppression of suffering being the dominant mechanism, but for other drugs, including nicotine and psychomotor stimulants such as cocaine and methamphetamine, increased postsynaptic dopamine receptor activation or increased levels of synaptic dopamine appear to be the primary factor. When people addicted to stimulants go through withdrawal, they do not experience the physical suffering associated with withdrawal from alcohol or opiates; instead they experience apathy, boredom, restlessness, and most importantly an overwhelming urge to consume more of the drug.[medical citation needed]
The addiction potential for stimulants is strongly dependent on the level of dopamine increase they produce.[medical citation needed]
Treatment of stimulant addiction is very difficult, because even if consumption ceases, the "craving" that comes with psychological withdrawal does not. Even still, when the craving seems to be extinct, it may reemerge when the individual experiences environmental stiumli (friends, locations, situations, etc.) that are associated with the drug. The brain mechanisms underlying these cravings have been a topic of extensive research. There is evidence that they are associated with long-lasting changes in the density of dopamine receptors in parts of the brain.[medical citation needed]
Pain
Dopamine has been demonstrated to play a role in pain processing in multiple levels of the central nervous system including the spinal cord, periaqueductal gray (PAG), thalamus, basal ganglia, and cingulate cortex. Accordingly, decreased levels of dopamine have been associated with painful symptoms that frequently occur in Parkinson's disease. Abnormalities in dopaminergic neurotransmission have also been demonstrated in painful clinical conditions, including burning mouth syndrome,[34] fibromyalgia, and restless legs syndrome. In general, the analgesic capacity of dopamine occurs as a result of dopamine D2 receptor activation; however, exceptions to this exist in the PAG, in which dopamine D1 receptor activation attenuates pain presumably via activation of neurons involved in descending inhibition.[35] In addition, D1 receptor activation in the insular cortex appears to attenuate subsequent pain-related behavior.
Nausea
Nausea and vomiting are largely determined by activity in a brainstem area known as the chemoreceptor trigger zone. This area contains a large population of type D2 dopamine receptors. Consequently, drugs that activate D2 receptors have a high potential to cause nausea. This group includes some medications that are administered for Parkinson's disease, as well as other dopamine agonists such as apomorphine. In many cases, D2-receptor antagonists such as metoclopramide are useful as anti-nausea drugs.
Psychosis
Abnormally high dopaminergic transmission has been linked to psychosis and schizophrenia.[36] However, clinical studies relating schizophrenia to brain dopamine metabolism have ranged from controversial to negative, with HVA levels in the CSF the same for schizophrenics and controls.[37] Increased dopaminergic functional activity, specifically in the mesolimbic pathway, is found in schizophrenic individuals. However, decreased activity in another dopaminergic pathway, the mesocortical pathway, may also be involved. The two pathways are thought to be responsible for differing sets of symptoms seen in schizophrenia.[citation needed]
Antipsychotic medications act largely as dopamine antagonists, inhibiting dopamine at the receptor level, and thereby blocking the effects of the neurochemical in a dose-dependent manner. The older, so-called typical antipsychotics most commonly act on D2 receptors,[38] while the atypical drugs also act on D1, D3 and D4 receptors, though they have a lower affinity for dopamine receptors in general.[39][40] The finding that drugs such as amphetamines, methamphetamine and cocaine, which can increase dopamine levels by more than tenfold,[41] can temporarily cause psychosis, provides further evidence for this link.[42] However, many non-dopaminergic drugs can induce acute and chronic psychosis.[43] The NMDA antagonists Ketamine and PCP both are used in research to reproduce the positive and negative symptoms commonly associated with schizophrenia.[44][45]
Dopaminergic dysregulation has also been linked to depressive disorders.[46] Early research in humans used various methods of analyzing dopamine levels and function in depressed patients. Studies have reported that there is decreased concentration of tyrosine, a precursor to dopamine, in the blood plasma, ventricular spinal fluid, and lumbar spinal fluid of depressed patients compared to control subjects.[47][48] One study measured the amount of homovanillic acid, the major metabolite of dopamine in the CSF, as a marker for the dopamine pathway turnover rate, and found decreased concentrations of homovanillic acid in the CSF of depressed patients.[49] Postmordem real time reverse transcriptase-polymerase chain reaction (RT-PCR) has also been used to find that gene expression of a specific subtype of dopamine receptor was elevated in the amygdala of people suffering from depression as compared to control subjects.[50]
The action of commonly used antidepressant drugs also has yielded information about possible alterations of the dopaminergic pathway in treating depression. It has been reported that many antidepressant drugs increase extracellular dopamine concentrations in the rat prefrontal cortex,[51] but vary greatly in their effects on the striatum and nucleus accumbens.[52][53] This can be compared to electro convulsive shock treatment (ECT), which has been shown to have a multiple fold increase in striatal dopamine levels in rats.[54]
More recent research studies with rodents have found that depression-related behaviors are associated with dopaminergic system dysregulation.[55] In rodents exposed to chronic mild stress, decreased escape behavior and decreased forced swimming is reversed with activation of the dopaminergic mesolimbic pathway.[55] Also, rodents that are susceptible to depression-related behavior after social defeat can have their behavior reversed with dopamine pathway activation.[56] Depletion of dopamine in the caudate nucleus and nucleus accumbens has also been reported in cases of learned helplessness in animals. These symptoms can be reversed with dopamine agonists and antidepressant administration prior to the learned helplessness protocol.[57]
Comparative biology and evolution
Microorganisms
There are no reports of dopamine in archaea, but it has been detected in some types of bacteria and in a type of protozoan called Tetrahymena.[58] Perhaps more importantly, there are types of bacteria that contain homologs of all the enzymes that animals use to synthesize dopamine. It has even been proposed that animals derived their dopamine-synthesizing machinery from bacteria, via horizontal gene transfer that may have occurred relatively late in evolutionary time, perhaps as a result of the symbiotic incorporation of bacteria into eukaryotic cells that gave rise to mitochondria.[59]
Animals
Dopamine is used as an intercellular messenger in virtually all multicellular animals. In sponges only a single report exists of the presence of dopamine, with no indication of its function;[60] however, dopamine has been reported in the nervous systems of numerous radially symmetric species, including cnidaria (jellyfish, hydra, corals, etc.).[61] This dates the emergence of dopamine as a neurotransmitter back to the earliest appearance of the nervous system, over 500 million years ago in the Cambrian era. Among existing species, dopamine functions as a neurotransmitter in vertebrates, echinoderms, arthropods, molluscs, and several types of worms.[62][63]
In every type of animal that has been examined, dopamine acts to modify motor behavior.[64] In the much-studied nematode worm Caenorhabditis elegans, it reduces locomotion and increases food-exploratory movements; in planarian worms it produces "screw-like" movements; in leeches it inhibits swimming and promotes crawling; etc. Across a wide range of vertebrates, dopamine has an "activating" effect on behavior-switching and response selection, comparable to its effect in mammals.[64]
Dopamine also consistently plays a role in reward learning, in all animal groups that have been examined except arthropods. In nematodes, planarians, molluscs, and vertebrates, animals can be trained to repeat an action if it is consistently followed by an increase in dopamine levels.[64] Arthropods are an exception, though. In these species — insects, crustaceans, etc. — dopamine has an aversive effect, and reward is instead mediated by octopamine, a neurotransmitter that is not found in vertebrates but is thought to be closely related to norepinephrine. In insects, dopamine increases aversion learning for olfactory stimuli as well as visual stimuli, and reduces approach learning for stimuli that are followed by rewards. It also improves recall for aversive memories and reduces recall for positive memories.[64] The origin of the striking reversal between dopamine's effects in arthropods versus all other types of animals has not been explained.
Plants

Many plants synthesize dopamine to varying degrees, including a variety of food plants. The highest concentrations have been observed in bananas — the fruit pulp of red and yellow bananas contains dopamine at levels of 40 to 50 parts per million by weight. Potatoes, avocados, broccoli, and Brussels sprouts may also contain dopamine at levels of 1 part per million or more; oranges, tomatoes, spinach, beans, and other plants contain measurable concentrations less than 1 part per million.[65] The dopamine in plants is synthesized from the amino acid tyrosine, by biochemical mechanisms similar to those that animals use. It can be metabolized in a number of ways, producing melanin and a variety of alkaloids as byproducts.[65] The functions of plant catecholamines have not been clearly established, but there is evidence that they play a role in the response to stressors such as bacterial infection, act as growth-promoting factors in some situations, and modify the way that sugars are metabolized. The receptors that mediate these actions have not yet been identified, nor have the intracellular mechanisms that they activate.[65]
Dopamine consumed in food cannot act on the brain, because it cannot cross the blood–brain barrier. However, there are also a variety of plants that contain L-DOPA, the metabolic precursor of dopamine.[66] The highest concentrations are found in the leaves and bean pods of plants of the genus Mucuna, especially in Mucuna pruriens (velvet beans), which have been used as a source for L-DOPA as a drug.[67] Another plant containing substantial amounts of L-DOPA is Vicia faba, the plant that produces fava beans (also known as "broad beans"). The level of L-DOPA in the beans, however, is much lower than in the pod shells and other parts of the plant.[68] The seeds of Cassia and Bauhinia trees also contain substantial amounts of L-DOPA.[66]
In the marine green alga Ulvaria obscura, which is a major component of some algal blooms, dopamine is present in very high concentrations, estimated at 4.4% of dry weight. There is evidence that this dopamine functions as an anti-herbivore defense, reducing consumption by snails and isopods.[69]
As a precursor for melanin
Melanins are a family of dark-pigmented substances that are found in a wide range of organisms. Their physical properties make them difficult to work with experimentally, and consequently a number of aspects of their biochemistry are not well understood. Chemically they are closely related to dopamine, and there is a type of melanin, known as "dopamine-melanin", that can be synthesized by oxidation of dopamine via the enzyme tyrosinase.[70] The melanin that darkens human skin is not of this type: it is synthesized by a pathway that uses L-DOPA as a precursor but not dopamine. However, there is substantial evidence that the "neuromelanin" that gives a dark color to the brain's substantia nigra is at least in part dopamine-melanin.[71]
Dopamine-derived melanin probably appears in at least some other biological systems as well. Some of the dopamine in plants is likely to be used as a precursor for dopamine-melanin.[72] The complex patterns that appear on butterfly wings, as well as black-and-white stripes on the bodies of insect larvae, are also thought to be caused by spatially structured accumulations of dopamine-melanin.[73]
Pharmacology
Dopamine as an injectable drug
Under the trade names Intropin, Dopastat, Revimine, or other names, dopamine can be used as a drug in injectable form. It is most commonly used in the treatment of severe hypotension, bradycardia (slow heart rate), circulatory shock, or cardiac arrest, especially in newborn infants. Its effects, depending on dosage, include an increase in sodium excretion by the kidneys, an increase in urine output, an increase in heart rate, and an increase in blood pressure. At a "cardiac dose" of 5 to 10 μg/kg/min, dopamine acts through the sympathetic nervous system to increase heart muscle contraction force and heart rate, thereby increasing cardiac output and blood pressure. At a "pressor dose" of 10 to 20 μg/kg/min, dopamine also causes vasoconstriction that further increases blood pressure, but can produce negative side effects such as an impairment of kidney function and cardiac arrhythmias.[74][75] Older literature also describes a so-called "renal dose" of 2 to 5 μg/kg/min thought to improve kidney function without other consequences, but recent reviews have concluded that doses at this low level are not clinically effective and may sometimes be harmful.[76]
L-DOPA
Levodopa is a dopamine precursor used in various forms to treat Parkinson's disease and dopa-responsive dystonia. It is typically co-administered with an inhibitor of peripheral decarboxylation (DDC, dopa decarboxylase), such as carbidopa or benserazide. Inhibitors of alternative metabolic route for dopamine by catechol-O-methyl transferase are also used. These include entacapone and tolcapone.
Psychomotor stimulants
Cocaine and amphetamines inhibit the re-uptake of dopamine; however, they influence separate mechanisms of action. Cocaine is a dopamine transporter and norepinephrine transporter blocker that competitively inhibits dopamine uptake to increase the lifetime of dopamine and augments an overabundance of dopamine (an increase of up to 150 percent) within the parameters of the dopamine neurotransmitters. Like cocaine, amphetamines increase the concentration of dopamine in the synaptic gap, but by a different mechanism. Amphetamine and methamphetamine are similar in structure to dopamine, and so can enter the terminal bouton of the presynaptic neuron via its dopamine transporters as well as by diffusing through the neural membrane directly.[77] Upon entering the presynaptic neuron, amphetamines activate Trace Amine Receptor 1 which, through protein kinase A and protein kinase C signaling, induces dopamine efflux and non-competitive reuptake inhibition.[77] Since amphetamines are structurally similar to trace amines, they are also substrates for monoamine transporters; consequently, they competitively inhibit the reuptake of dopamine and other monoamines as well.[77]
Antipsychotic drugs
A range of drugs that reduce dopamine activity have been found useful in the treatment of schizophrenia and other disorders that produce psychosis. These antipsychotic drugs are also sometimes known as neuroleptics or "major tranquilizers", in contrast to "minor tranquilizers" such as Valium that are used to treat anxiety or sleep disorders. These drugs have a broadly suppressive effect on most types of active behavior, and particularly reduce the delusional and agitated behavior characteristic of overt psychosis. The introduction of the first widely used antipsychotic drug, chlorpromazine (Thorazine), in the 1950s, led to the release of many schizophrenia patients from institutions in the years that followed.
Even so, the widespread use of antipsychotic drugs has long been controversial. There are several reasons for this. First, these drugs are perceived as very aversive by people who have to take them, because they produce a general dullness of thought and suppress the ability to experience pleasure.[78] Second, it is difficult to show that they act specifically against psychotic behaviors rather than merely suppressing all types of active behavior. Third, they can produce a range of serious side effects, including weight gain, diabetes, fatigue, sexual dysfunction, hormonal changes, and a type of movement disorder known as tardive dyskinesia. Some of these side effects may continue long after the cessation of drug use, or even permanently.
The first drugs introduced specifically for the treatment of psychosis all had strong direct effects on multiple aspects of dopamine function. Drugs of this type are known as "typical antipsychotics". Because of the problems they cause, there has been wide interest in newer types of drugs known as "atypical antipsychotics" or "second-generation antipsychotics", which aim to target the specific types of dopamine receptors involved in psychosis, and thereby reduce psychotic symptoms without producing as many undesirable side effects. There remains substantial dispute, however, about how much of an improvement in the patient experience these drugs produce.
Toxicity
The LD50, or dose which is expected to be lethal in 50% of the population, has been found to be: 59 mg/kg (mouse; administered i.v.); 950 mg/kg (mouse; administered i.p.); 163 mg/kg (rat; administered i.p.); 79 mg/kg (dog; administered i.v.)[79]
Binding profile of dopamine[80]
| Macromolecule | Ki (nM) |
|---|---|
| 5-HT1A | 8248 |
| 5-HT7 | >10000 |
| D1 | 130 |
| D2 | 598 |
| D3 | 32.5 |
| D4 | 182.6 |
| D5 | 228 |
| DAT | 67 |
| NET | 323 |
| TAAR1 | 422 |
Biochemical mechanisms
Structurally, dopamine belongs to the catecholamine and phenethylamine classes. In biological systems, dopamine is synthesized in brain cells and adrenal cells from the precursor L-DOPA. In brain cells, it is transported to synaptic sites and packaged into vesicles for release, which occurs during synaptic transmission. After release, free dopamine is either reabsorbed into the presynaptic terminal for reuse, or broken down by the enzymes monoamine oxidase or COMT, producing a variety of degradation metabolites.
Biosynthesis
Dopamine is synthesized in a restricted set of cell types, mainly neurons and cells in the medulla of the adrenal glands. This is the metabolic pathway:
- L-Phenylalanine → L-Tyrosine → L-DOPA → Dopamine
Thus the direct precursor of dopamine is L-DOPA, but this itself can be synthesized from the essential amino acid phenylalanine or the non-essential amino acid tyrosine. These amino acids are found in nearly every protein and as such are provided from ingestion of protein-containing food, with tyrosine being the most common. Although dopamine itself is also found in many types of food, it is incapable of crossing the blood–brain barrier that surrounds and protects the brain. It must therefore be synthesized inside the brain in order to perform its neural actions.
L-Phenylalanine is converted into L-tyrosine by the enzyme tyrosine hydroxylase (TH) (also known as phenylalanine hydroxylase), with molecular oxygen (O2) and tetrahydrobiopterin (THB) as cofactors. L-Tyrosine is converted into L-DOPA by the enzyme tyrosine hydroxylase (TH), with tetrahydrobiopterin (THB), O2, and ferrous iron (Fe2+) as cofactors. L-DOPA is converted into dopamine by the enzyme aromatic L-amino acid decarboxylase (AAAD; also known as DOPA decarboxylase (DDC)), with pyridoxal phosphate (PLP) as the cofactor.
Dopamine itself is also used as precursor in the synthesis of the neurotransmitters norepinephrine and epinephrine. Dopamine is converted into norepinephrine by the enzyme dopamine β-hydroxylase (DBH), with O2 and L-ascorbic acid as cofactors. Norepinephrine is converted into epinephrine by the enzyme phenylethanolamine N-methyltransferase (PNMT) with S-adenosyl-L-methionine (SAMe) as the cofactor.
It should be noted that some of the cofactors also require their own synthesis. Deficiency in any required amino acid or cofactor will result in subsequent dopamine, norepinephrine, and epinephrine biosynthesis impairment and deficiency.

DBH: Dopamine β-hydroxylase;
AADC:Aromatic L-amino acid decarboxylase;
AAAH: (Biopterin-dependent) aromatic amino acid hydroxylase;
COMT: Catechol O-methyltransferase;
PNMT: Phenylethanolamine N-methyltransferase
Storage, release, and reuptake
Inside the brain dopamine functions as a neurotransmitter, and is controlled by a set of mechanisms that are common to all neurotransmitters. After synthesis, dopamine is transported from the cytosol[citation needed] into synaptic vesicles by the vesicular monoamine transporter 2 (VMAT2). Dopamine is stored in and remains in these vesicles until an action potential occurs and causes the contents of the vesicles to be ejected into the synaptic cleft.
Once in the synapse, dopamine binds to and activates dopamine receptors, which can be located either on postsynaptic target cells or on the membrane of the dopamine-releasing cell itself (i.e., autoreceptors).
After an action potential, the dopamine molecules quickly become unbound from their receptors. They are then absorbed back into the presynaptic cell, via reuptake mediated either by the high-affinity dopamine transporter (DAT) or by the low-affinity plasma membrane monoamine transporter (PMAT). Once back in the cytosol, dopamine is subsequently repackaged into vesicles by VMAT2, making it available for future release.
Degradation
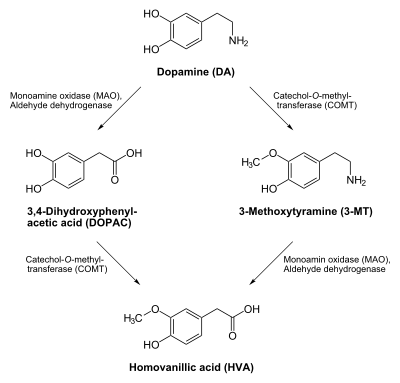
Dopamine is broken down into inactive metabolites by a set of enzymes, monoamine oxidase (MAO), aldehyde dehydrogenase (ALDH), and catechol-O-methyl transferase (COMT), acting in sequence. Both isoforms of MAO, MAO-A and MAO-B, are equally effective.
The metabolites produced by these processes are:
- DOPAL (3,4-Dihydroxyphenylacetaldehyde)
- DOPAC (3,4-Dihydroxyphenylacetic acid)
- DOPET (3,4-dihydroxyphenylethanol, also known as Hydroxytyrosol)
- MOPET (3-methoxy-4-hydroxyphenylethanol, also known as Homovanillyl alcohol)
- 3-MT (3-Methoxytyramine)
- HVA (Homovanillic acid)
All of these are intermediate metabolites except MOPET and HVA, which are filtered from the bloodstream by the kidneys and then excreted in the urine.
The specific reactions that make up these pathways are:
- Dopamine → DOPAL, mediated by MAO
- DOPAL → DOPAC, mediated by ALDH
- DOPAL → DOPET, mediated by aldose reductase (minor pathway)
- DOPAC → HVA, mediated by COMT
- DOPET → MOPET, mediated by COMT
- Dopamine → 3-MT, mediated by COMT
- 3-MT → HVA, mediated by MAO
In most areas of the brain, including the striatum and basal ganglia, dopamine is inactivated by reuptake via the DAT, then enzymatic breakdown by MAO into DOPAC. In the prefrontal cortex, however, there are very few DAT proteins, and dopamine is inactivated instead by reuptake via the norepinephrine transporter (NET), presumably on neighboring norepinephrine neurons, then enzymatic breakdown by COMT into 3-MT.[81] The DAT pathway is roughly an order of magnitude faster than the NET pathway: in mice, dopamine concentrations decay with a half-life of 200 milliseconds in the caudate nucleus (which uses the DAT pathway) versus 2,000 milliseconds in the prefrontal cortex.[82] Dopamine that is not broken down by enzymes is repackaged into vesicles for future release.
Chemistry


Chemically, a dopamine molecule consists of a catechol structure (a benzene ring with two hydroxyl side groups) with one amine group attached. As such, dopamine is the simplest possible catecholamine, a family that also includes the neurotransmitters norepinephrine and epinephrine. The presence of a benzene ring with an attached amine group makes it a phenethylamine, a family that includes numerous psychoactive drugs.
Dopamine, like most amines, is an organic base. At neutral or acidic pH levels it is generally protonated. The protonated form is highly water-soluble and relatively stable, though it is capable of oxidizing if exposed to oxygen or other oxidants. At basic pH levels, dopamine becomes deprotonated. In this free base form it is less soluble and also highly reactive and easily oxidized. Because of this pH-dependence, dopamine is supplied for chemical or pharmaceutical use in the form of dopamine hydochloride, that is, the hydrochloride salt that is created when dopamine is combined with hydrochloric acid. In dry form, dopamine hydrochloride is a fine colorless powder. When dissolved in distilled water it gives a solution that is mildly acidic and therefore relatively stable. It cannot, however, be combined with alkaline solutions such as a bicarbonate buffer without being rendered inactive.
Oxidation
Dopamine in the body is normally broken down by oxidation catalyzed by the enzyme monoamine oxidase. However, dopamine is also capable of autoxidation, that is, direct reaction with oxygen, yielding quinones plus various free radicals as products.[83] The rate of autoxidation can be increased by the presence of ferrous iron or other factors. The ability of dopamine autoxidation to produce quinones and free radicals makes it a potent cell toxin, and there is evidence that this mechanism may contribute to cell loss that occurs in Parkinson's disease or other conditions.[84]
Polydopamine
Research motivated by mussel adhesive proteins led to the discovery in 2007 that a wide variety of materials, if placed in a solution of dopamine at slightly basic pH, will become coated with a layer of polymerized dopamine, often referred to as polydopamine.[85][86] This polymerized dopamine forms by a spontaneous oxidation reaction, and is formally a type of melanin.[87] Synthesis usually involves reaction of dopamine hydrochloride with Tris as a base in water. The structure of polydopamine is unknown.[86]
Polydopamine coatings can form on objects ranging in size from nanoparticles to large surfaces. Polydopamine layers have chemical properties that have the potential to be extremely useful, and numerous studies have examined their possible applications. At the simplest level, they can be used for protection against damage by light, or to form capsules for drug delivery. At a more sophisticated level, their adhesive properties may make them useful as substrates for biosensors or other biologically active macromolecules.[87]
History
Dopamine was first synthesized in 1910 by George Barger and James Ewens at Wellcome Laboratories in London, England.[88] It was named dopamine because it is a monoamine whose precursor in the Barger-Ewens synthesis is 3,4-dihydroxyphenylalanine (levodopamine or L-DOPA). Dopamine's function as a neurotransmitter was first recognized in 1958 by Arvid Carlsson and Nils-Åke Hillarp at the Laboratory for Chemical Pharmacology of the National Heart Institute of Sweden.[89] Carlsson was awarded the 2000 Nobel Prize in Physiology or Medicine for showing that dopamine is not only a precursor of norepinephrine (noradrenaline) and epinephrine (adrenaline), but also a neurotransmitter.
See also
- Addiction
- Amphetamine
- Antipsychotic
- Catecholamine
- Catechol-O-methyl transferase
- Classical conditioning
- Cocaine
- Depression
- Dopamine hypothesis of schizophrenia
- Dopamine reuptake inhibitor
- Epinine (N-methyldopamine)
- Limbic system
- Methylphenidate
- N,N-Dimethyldopamine
- Neurotransmitter
- Operant conditioning
- Parkinson's disease
- Prolactinoma
- Schizophrenia
- Selegiline
- Serotonin
References
- ↑ Schultz W (2007). "Multiple dopamine functions at different time courses". Annu. Rev. Neurosci. 30: 259–88. doi:10.1146/annurev.neuro.28.061604.135722. PMID 17600522.
- ↑ A. Dahlström and K. Fuxe (1964). "Evidence for the existence of monoamine-containing neurons in the central nervous system. I. Demonstration of monoamines in the cell bodies of brain stem neurons". Acta physiologica Scandinavica. Supplementum 232: 1–55. PMID 14229500.
- ↑ 3.0 3.1 Björklund A, Dunnett SB (May 2007). "Dopamine neuron systems in the brain: an update". Trends Neurosci. 30 (5): 194–202. doi:10.1016/j.tins.2007.03.006. PMID 17408759.
- ↑ Christine CW, Aminoff MJ (September 2004). "Clinical differentiation of parkinsonian syndromes: prognostic and therapeutic relevance". Am. J. Med. 117 (6): 412–9. doi:10.1016/j.amjmed.2004.03.032. PMID 15380498.
- ↑ DeLong M, Wichmann T (April 2010). "Changing views of basal ganglia circuits and circuit disorders". Clin EEG Neurosci 41 (2): 61–7. doi:10.1177/155005941004100204. PMID 20521487.
- ↑ Robbins TW, Arnsten AF. (2009). "The neuropsychopharmacology of fronto-executive function: monoaminergic modulation". Annu Rev Neurosci. 32: 267–87. doi:10.1146/annurev.neuro.051508.135535. PMC 2863127. PMID 19555290.
- ↑ Paulus W, Schomburg ED (June 2006). "Dopamine and the spinal cord in restless legs syndrome: does spinal cord physiology reveal a basis for augmentation?". Sleep Med Rev 10 (3): 185–96. doi:10.1016/j.smrv.2006.01.004. PMID 16762808.
- ↑ 8.0 8.1 Ben-Jonathan N, Hnasko R (2001). "Dopamine as a Prolactin (PRL) Inhibitor" (PDF). Endocrine Reviews 22 (6): 724–763. doi:10.1210/er.22.6.724. PMID 11739329.
- ↑ Witkovsky P (January 2004). "Dopamine and retinal function". Doc Ophthalmol 108 (1): 17–40. doi:10.1023/B:DOOP.0000019487.88486.0a. PMID 15104164.
- ↑ 10.0 10.1 10.2 Eisenhofer G, Kopin IJ, Goldstein DS (September 2004). "Catecholamine metabolism: a contemporary view with implications for physiology and medicine". Pharmacol. Rev. 56 (3): 331–49. doi:10.1124/pr.56.3.1. PMID 15317907.
- ↑ Missale, C; Nash, SR; Robinson, SW; Jaber, M; Caron, MG (1998). "Dopamine receptors: from structure to function". Physiological reviews 78 (1): 189–225. PMID 9457173.
- ↑ 12.0 12.1 Buttarelli FR, Fanciulli A, Pellicano C, Pontieri FE (June 2011). "The dopaminergic system in peripheral blood lymphocytes: from physiology to pharmacology and potential applications to neuropsychiatric disorders". Curr Neuropharmacol 9 (2): 278–88. doi:10.2174/157015911795596612. PMC 3131719. PMID 22131937.
- ↑ Basu, S; Dasgupta, PS. (2000). "Dopamine, a neurotransmitter, influences the immune system". J Neuroimmunol 102 (2): 113–24. doi:10.1016/S0165-5728(99)00176-9. PMID 10636479.
- ↑ Sarkar, C; Basu, B; Chakroborty, D; Dasgupta, PS; Basu, S (2010). "The immunoregulatory role of dopamine: an update". Brain, behavior, and immunity 24 (4): 525–8. doi:10.1016/j.bbi.2009.10.015. PMC 2856781. PMID 19896530.
- ↑ Carey RM (September 2001). "Theodore Cooper Lecture: Renal dopamine system: paracrine regulator of sodium homeostasis and blood pressure". Hypertension 38 (3): 297–302. doi:10.1161/hy0901.096422. PMID 11566894.
- ↑ 16.0 16.1 Rubí B, Maechler P (December 2010). "Minireview: new roles for peripheral dopamine on metabolic control and tumor growth: let's seek the balance". Endocrinology 151 (12): 5570–81. doi:10.1210/en.2010-0745. PMID 21047943.
- ↑ Grace AA, (1991). "Phasic versus tonic dopamine release and the modulation of dopamine system responsivity: A hypothesis for the eitiology of schizophrenia". Neuroscience 41 (1): 1–24. doi:10.1016/0306-4522(91)90196-U. PMID 1676137.
- ↑ Grace AA, Bunney BS (1984). "The control of firing pattern in nigral dopamine neurons: single spike firing" (PDF). Journal of Neuroscience 4 (11): 2866–2876. PMID 6150070.
- ↑ Grace AA, Bunney BS (1984). "The control of firing pattern in nigral dopamine neurons: burst firing" (PDF). Journal of Neuroscience 4 (11): 28677–2890. PMID 6150071.
- ↑ Calabresi P, Picconi B, Tozzi A, Di Filippo M (May 2007). "Dopamine-mediated regulation of corticostriatal synaptic plasticity". Trends Neurosci. 30 (5): 211–9. doi:10.1016/j.tins.2007.03.001. PMID 17367873.
- ↑ 21.0 21.1 Arias-Carrión O, Pöppel E (2007). "Dopamine, learning and reward-seeking behavior". Act Neurobiol Exp 67 (4): 481–488.
- ↑ 22.0 22.1 Wise RA (1996). "Addictive drugs and brain stimulation reward". Annu. Rev. Neurosci. 19: 319–40. doi:10.1146/annurev.ne.19.030196.001535. PMID 8833446.
- ↑ 23.0 23.1 Schultz W (2002). "Getting formal with dopamine and reward". Neuron 36 (2): 241–263. doi:10.1016/S0896-6273(02)00967-4. PMID 12383780.
- ↑ 24.0 24.1 24.2 24.3 Matsumoto M, Hikosaka O. (2009). "Two types of dopamine neuron distinctly convey positive and negative motivational signals". Nature 459 (7248): 837–41. doi:10.1038/nature08028. PMC 2739096. PMID 19448610.
- ↑ Williams SM, Goldman-Rakic PS. (1998). "Widespread origin of the primate mesofrontal dopamine system". Cereb Cortex. 8 (4): 321–45. doi:10.1093/cercor/8.4.321. PMID 9651129.
- ↑ Deutch AY, Roth RH. (1990). "The determinants of stress-induced activation of the prefrontal cortical dopamine system". Prog Brain Res. Progress in Brain Research 85: 367–402. doi:10.1016/S0079-6123(08)62691-6. ISBN 9780444811240. PMID 2094906.
- ↑ Brozoski TJ, Brown RM, Rosvold HE, Goldman PS. (1979). "Cognitive deficit caused by regional depletion of dopamine in prefrontal cortex of rhesus monkey". Science 205 (4409): 929–32. Bibcode:1979Sci...205..929B. doi:10.1126/science.112679. PMID 112679.
- ↑ Arnsten AF, Wang MJ, Paspalas CD. (2012). "Neuromodulation of thought: flexibilities and vulnerabilities in prefrontal cortical network synapses". Neuron 76 (1): 223–39. doi:10.1016/j.neuron.2012.08.038. PMC 3488343. PMID 23040817.
- ↑ Vijayraghavan S, Wang M, Birnbaum SG, Williams GV, Arnsten AF. (2007). "Inverted-U dopamine D1 receptor actions on prefrontal neurons engaged in working memory". Nat Neurosci. 10 (3): 376–84. doi:10.1038/nn1846. PMID 17277774.
- ↑ Wang M, Vijayraghavan S, Goldman-Rakic PS. (2004). "Selective D2 receptor actions on the functional circuitry of working memory". Science 303 (5659): 853–6. Bibcode:2004Sci...303..853W. doi:10.1126/science.1091162. PMID 14764884.
- ↑ Merims D, Giladi N (2008). "Dopamine dysregulation syndrome, addiction and behavioral changes in Parkinson's disease". Parkinsonism & Related Disorders 14 (4): 273–80. doi:10.1016/j.parkreldis.2007.09.007. PMID 17988927.
- ↑ Wu J, Xiao H, Sun H, Zou L, Zhu LQ. (2012). "Role of dopamine receptors in ADHD: a systematic meta-analysis". Mol Neurobiol. 45: 605–20.
- ↑ Berridge CW, Devilbiss DM. (2011). "Psychostimulants as cognitive enhancers: the prefrontal cortex, catecholamines, and attention-deficit/hyperactivity disorder". Biol Psychiatry 69 (12): e101–11.
- ↑ Jääskeläinen, SK; Rinne, JO; Forssell, H; Tenovuo, O; Kaasinen, V; Sonninen, P; Bergman, J. (2001). "Role of the dopaminergic system in chronic pain -- a fluorodopa-PET study". Pain 90 (3): 257–60. doi:10.1016/S0304-3959(00)00409-7. PMID 11207397.
- ↑ Wood, PB. (2008). "Role of central dopamine in pain and analgesia". Expert Rev Neurother 8 (5): 781–97. doi:10.1586/14737175.8.5.781. PMID 18457535.
- ↑ "Disruption of gene interaction linked to schizophrenia". St. Jude Children's Research Hospital. Retrieved 6 July 2006.
- ↑ Maas, J.W.; Bowden CL, Miller AL, Javors MA, Funderburg LG, Berman N, Weintraub ST. (1997). "Schizophrenia, psychosis, and cerebral spinal fluid homovanillic acid concentrations". Schizophrenia Bulletin. 23 (1): 147–154. doi:10.1093/schbul/23.1.147. PMID 9050120.
- ↑ http://www.williams.edu/imput/synapse/pages/IIIB5.htm
- ↑ http://bjp.rcpsych.org/cgi/content/full/181/4/271
- ↑ Durcan, M; Rigdon, GC; Norman, MH; Morgan, PF (1995). "Is clozapine selective for the dopamine D4 receptor?". Life Sciences 57 (18): PL275–83. doi:10.1016/0024-3205(95)02151-8. PMID 7475902.
- ↑ Methamphetamine 101
- ↑ Lieberman, J.A.; JM Kane, J. Alvir (1997). "Provocative tests with psychostimulant drugs in schizophrenia". Psychopharmacology (Berl). 91 (4): 415–433. doi:10.1007/BF00216006. PMID 2884687.
- ↑ Cardinal, R.N. & Bullmore, E.T., The Diagnosis of Psychosis, Cambridge University Press, 2011, ISBN 978-0-521-16484-9
- ↑ Jentsch, J.; Roth, R. H. (1999). "The Neuropsychopharmacology of Phencyclidine from NMDA Receptor Hypofunction to the Dopamine Hypothesis of Schizophrenia". Neuropsychopharmacology 20 (3): 201–225. doi:10.1016/S0893-133X(98)00060-8. PMID 10063482.
- ↑ Abi-Saab, WM; D'Souza DC, Moghaddam B, Krystal JH (1998). "The NMDA antagonist model for schizophrenia: promise and pitfalls". Pharmacopsychiatry 31 (2): 104–109. doi:10.1055/s-2007-979354. PMID 9754841.
- ↑ Galani, VJ; Rana DG (2011). "Depression and antidepressants with dopamine hypothesis-A review". IJPFR 1 (2): 45–60.
- ↑ Denkert, O; Renz A, Marano C, Matussek N. (1971). "Altered tyrosine daytime plasma levels in edogenous depressed patients". Arch gen Psychiat 25 (4): 359–363. doi:10.1001/archpsyc.1971.01750160071013. PMID 5116991.
- ↑ Birkmayer, W; Linauer W, Storung D (1970). "Tyrosin and tryptophan- metabolisms in depression patients". Arch Psychiar Nervenkr 213 (4): 377–387. doi:10.1007/BF00341554.
- ↑ Bowers, MB; Heninger GR, Gerbode F. (1969). "Cerebrospinal fluid 5-hydroxyindoleacetic acid and homovanillic acid in psychiatric patients". Int J Neuropharmacol 8 (3): 255–262. doi:10.1016/0028-3908(69)90046-X. PMID 5796265.
- ↑ Lianbin, X; Katalin S, Attila S, Violetta K, Craig A, Stockmeier C, Beata K, John K, Gregory A, Ordwaya (2008). "Dopamine receptor gene expression in human amygdaloid nuclei: Elevated D4 receptor mRNA in major depression". Brain Res 1207: 214–224. doi:10.1016/j.brainres.2008.02.009. PMC 2577810. PMID 18371940.
- ↑ Carlson, JN; Visker KE, Nielsen DM, Keller RW, Glick SD (1996). "Chronic antidepressant drug treatment reduces turning behavior and increases dopamine levels in the medial prefrontal cortex". Brain Res 707 (1): 122–126. doi:10.1016/0006-8993(95)01341-5. PMID 8866721.
- ↑ Ainsworth, K; Smith SE, Zetterstrom TS, Pei Q, Franklin M, Sharp T (1998). "Effect of antidepressant drugs on dopamine D1 and D2 receptor expression and dopamine release in the nucleus accumbens of the rat". Psychopharmacology 140 (4): 470–477. doi:10.1007/s002130050791. PMID 9888623.
- ↑ Meltzer, TL; Wiley JN, Williams AE, Heffner TG (1988). "Evidence for postsynaptic dopamine effects of B-HT 920 in the presence of the dopamine D1 agonist SKF 38393". Psychopharmacology 95 (3): 329–332. doi:10.1007/BF00181942. PMID 2901126.
- ↑ Nomikos, GG; Damsma G, Wenkstern D, Fibiger HC (1989). "Acute effects of bupripion on extracellular dopamine concentration in rat striatum and nucleus accumbens studies by in vivo microdialysis study". Neuropsychopharmacol 4: 65–69.
- ↑ 55.0 55.1 Deisseroth, Julie J.; Mirzabekov, Warden, Ferenczi, Tsai, Finkelstein, kim, Adhikari, Thompson, Andalman, Gunaydin, Witten & Deisseroth (2012). "Dopamine neurons modulate neural encoding and expression of depression-related behaviour". Nature 493 (7433): 537–541. Bibcode:2013Natur.493..537T. doi:10.1038/nature11740. PMID 23235822.
- ↑ Han, D; Walsh, Friedman, Juarez, Ku, Koo, Ferguson, Tsai, Pomeranz, Christoffel, Nectow, Ekstrand, Domingos, Mazei-Robison, Mouzon, Lobo, Neve, Friedman, Russo, Deisseroth, Nestler, Han (2013). "Rapid regulation of depression-related behaviors by control of midbrain dopamine neurons". Nature 493 (7433): 532–536. Bibcode:2013Natur.493..532C. doi:10.1038/nature11713. PMC 3554860. PMID 23235832.
- ↑ Muscat, R; Sampson D, Willner P. (1990). "Dopaminergic mechanisms of imipramine action in an animal model of depression". Biol Psychiatry 28 (3): 223–230. doi:10.1016/0006-3223(90)90577-O. PMID 2378927.
- ↑ Roshchina VV (2010). "Evolutionary considerations of neurotransmitters in microbial, plant, and animal cells". In Lyte M, Primrose PEPE. Microbial Endocrinology. New York: Springer. pp. 17–52. ISBN 978-1-4419-5576-0.
- ↑ Iyer LM, Aravind L, Coon SL, Klein DC, Koonin EV (July 2004). "Evolution of cell-cell signaling in animals: did late horizontal gene transfer from bacteria have a role?". Trends Genet. 20 (7): 292–9. doi:10.1016/j.tig.2004.05.007. PMID 15219393.
- ↑ Liu H, Mishima Y, Fujiwara T, Nagai H, Kitazawa A, Mine Y, et al. (2004). "Isolation of Araguspongine M, a new stereoisomer of an Araguspongine/Xestospongin alkaloid, and dopamine from the marine sponge Neopetrosia exigua collected in Palau". Marine Drugs 2 (4): 154–163. doi:10.3390/md204154.
- ↑ Kass-Simon G, Pierobon P (January 2007). "Cnidarian chemical neurotransmission, an updated overview". Comp. Biochem. Physiol., Part a Mol. Integr. Physiol. 146 (1): 9–25. doi:10.1016/j.cbpa.2006.09.008. PMID 17101286.
- ↑ Cottrell GA (January 1967). "Occurrence of dopamine and noradrenaline in the nervous tissue of some invertebrate species". Br J Pharmacol Chemother 29 (1): 63–9. doi:10.1111/j.1476-5381.1967.tb01939.x. PMC 1557178. PMID 19108240.
- ↑ Kindt KS, Quast KB, Giles AC, et al. (August 2007). "Dopamine mediates context-dependent modulation of sensory plasticity in C. elegans". Neuron 55 (4): 662–76. doi:10.1016/j.neuron.2007.07.023. PMID 17698017.
- ↑ 64.0 64.1 64.2 64.3 Barron AB, Søvik E, Cornish JL (2010). "The roles of dopamine and related compounds in reward-seeking behavior across animal phyla". Front Behav Neurosci 4: 163. doi:10.3389/fnbeh.2010.00163. PMC 2967375. PMID 21048897.
- ↑ 65.0 65.1 65.2 Kulma A, Szopa J (2007). "Catecholamines are active compounds in plants". Plant Science 172 (3): 433–440. doi:10.1016/j.plantsci.2006.10.013.
- ↑ 66.0 66.1 Ingle PK (2003). "L-DOPA bearing plants" (PDF). Natural Product Radiance 2: 126–133. Retrieved 3 February 2014.
- ↑ Wichers HJ, Visser JF, Huizing HJ, Pras N (1993). "Occurrence of L-DOPA and dopamine in plants and cell cultures of Mucuna pruriens and effects of 2, 4-d and NaCl on these compounds". Plant Cell, Tissue and Organ Culture 33 (3): 259–264. doi:10.1007/BF02319010.
- ↑ Longo R (1974). "Distribution of L-dopa and related amino acid in Vicia". Phytochemistry 13: 167–171. doi:10.1016/S0031-9422(00)91287-1.
- ↑ Van Alstyne, Kathryn L.; Nelson, Amorah V.; Vyvyan, James R.; Cancilla, Devon A. (2006). "Dopamine functions as an antiherbivore defense in the temperate green alga Ulvaria obscura". Oecologia 148 (2): 304–311. doi:10.1007/s00442-006-0378-3. PMID 16489461.
- ↑ Simon JD, Peles D, Wakamatsu K, Ito S (October 2009). "Current challenges in understanding melanogenesis: bridging chemistry, biological control, morphology, and function". Pigment Cell Melanoma Res 22 (5): 563–79. doi:10.1111/j.1755-148X.2009.00610.x. PMID 19627559.
- ↑ Fedorow H, Tribl F, Halliday G, Gerlach M, Riederer P, Double KL (February 2005). "Neuromelanin in human dopamine neurons: comparison with peripheral melanins and relevance to Parkinson's disease". Prog. Neurobiol. 75 (2): 109–24. doi:10.1016/j.pneurobio.2005.02.001. PMID 15784302.
- ↑ Andrews RS, Pridham JB (1967). "Melanins from DOPA-containing plants". Phytochemistry 6 (1): 13–18. doi:10.1016/0031-9422(67)85002-7.
- ↑ Beldade P, Brakefield PM (June 2002). "The genetics and evo-devo of butterfly wing patterns". Nature Reviews Genetics 3 (6): 442–52. doi:10.1038/nrg818. PMID 12042771.
- ↑ Bronwen Jean Bryant; Kathleen Mary Knights (15 November 2009). Pharmacology for Health Professionals (2nd ed.). Elsevier Australia. p. 192. ISBN 978-0-7295-3929-6. Retrieved 9 June 2011.
- ↑ de Backer D, Biston P, Devriendt J, et al. (2010). "Comparison of Dopamine and Norepinephrine in the Treatment of Shock". New Engl. J. Med. 362 (9): 779–789. doi:10.1056/NEJMoa0907118. PMID 20200382.
- ↑ Karthik S, Lisbon A (2006). "Low-dose dopamine in the intensive care unit". Semin Dial 19 (6): 465–71. doi:10.1111/j.1525-139X.2006.00208.x. PMID 17150046.
- ↑ 77.0 77.1 77.2 Miller, GM (January 2011). "The emerging role of trace amine-associated receptor 1 in the functional regulation of monoamine transporters and dopaminergic activity". Journal of Neurochemistry 116 (2): 164–76. doi:10.1111/j.1471-4159.2010.07109.x. PMC 3005101. PMID 21073468.
- ↑ Lambert M, Schimmelmann B, Karow A, Naber D (2003). "Subjective well-being and initial dysphoric reaction under antipsychotic drugs - concepts, measurement and clinical relevance". Pharmacopsychiatry 36 (Suppl 3): S181–90. doi:10.1055/s-2003-45128. PMID 14677077.
- ↑ R. J. Lewis (Ed.) (2004), Sax's Dangerous Properties of Industrial Materials, 11th Ed., p. 1552, Wiley & Sons, Hoboken, NJ.
- ↑ Roth, BL; Driscol, J (12 January 2011). "PDSP Ki Database". Psychoactive Drug Screening Program (PDSP). University of North Carolina at Chapel Hill and the United States National Institute of Mental Health. Retrieved 15 November 2013.
- ↑ Morón, JA; Brockington, A; Wise, RA; Rocha, BA; Hope, BT (2002). "Dopamine uptake through the norepinephrine transporter in brain regions with low levels of the dopamine transporter: evidence from knock-out mouse lines". Journal of Neuroscience 22 (2): 389–95. PMID 11784783.
- ↑ Yavich, L; Forsberg, MM; Karayiorgou, M; Gogos, JA; Männistö, PT (2007). "Site-specific role of catechol-O-methyltransferase in dopamine overflow within prefrontal cortex and dorsal striatum". Journal of Neuroscience 27 (38): 10196–209. doi:10.1523/JNEUROSCI.0665-07.2007. PMID 17881525.
- ↑ Sulzer D, Zecca L (February 2000). "Intraneuronal dopamine-quinone synthesis: a review". Neurotox Res 1 (3): 181–95. doi:10.1007/BF03033289. PMID 12835101.
- ↑ Miyazaki I, Asanuma M (June 2008). "Dopaminergic neuron-specific oxidative stress caused by dopamine itself" (PDF). Acta Med. Okayama 62 (3): 141–50. PMID 18596830.
- ↑ Mussel-Inspired Surface Chemistry for Multifunctional Coatings Haeshin Lee, Shara M. Dellatore, William M. Miller, Phillip B. Messersmith Science 19 October 2007: Vol. 318 no. 5849 pp. 426–430 doi:10.1126/science.1147241
- ↑ 86.0 86.1 Perspectives on poly(dopamine) Daniel R. Dreyer, Daniel J. Miller, Benny D. Freeman, Donald R. Paul and Christopher W. Bielawski Chem" Sci 2013, Advance Article doi:10.1039/C3SC51501J
- ↑ 87.0 87.1 Lynge ME, van der Westen R, Postma A, Städler B (December 2011). "Polydopamine--a nature-inspired polymer coating for biomedical science". Nanoscale 3 (12): 4916–28. doi:10.1039/c1nr10969c. PMID 22024699.
- ↑ Fahn, Stanley, "The History of Levodopa as it Pertains to Parkinson's disease," Movement Disorder Society's 10th International Congress of Parkinson's Disease and Movement Disorders on November 1, 2006, in Kyoto, Japan.
- ↑ Benes, F.M. (2001). "Carlsson and the discovery of dopamine". Trends in Pharmacological Sciences 22 (1): 46–47. doi:10.1016/S0165-6147(00)01607-2. PMID 11165672.
External links
| Look up Dopamine in Wiktionary, the free dictionary. |
- DrugBank APRD00085
- U.S. National Library of Medicine: Drug Information Portal - Dopamine
- Dopamine: analyte monograph - The Association for Clinical Biochemistry and Laboratory Medicine
| |||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||
| ||||||||||||||||||