Biological neuron model
A biological neuron model (also known as spiking neuron model) is a mathematical description of the properties of nerve cells, or neurons, that is designed to accurately describe and predict biological processes. This is in contrast to the artificial neuron, which aims for computational effectiveness, although these goals sometimes overlap.
Artificial neuron abstraction
The most basic model of a neuron consists of an input with some synaptic weight vector and an activation function or transfer function inside the neuron determining output. This is the basic structure used in artificial neurons, which in a neural network often looks like
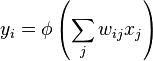
where yi is the output of the i th neuron, xj is the jth input neuron signal, wij is the synaptic weight (or strength of connection) between the neurons i and j, and φ is the activation function. Some of the earliest biological models took this form until kinetic models such as the Hodgkin-Huxley model became dominant.
Biological abstraction
In the case of modelling a biological neuron, physical analogues are used in place of abstractions such as "weight" and "transfer function". The input to a neuron is often described by an ion current through the cell membrane that occurs when neurotransmitters cause an activation of ion channels in the cell. We describe this by a physical time-dependent current I(t). The cell itself is bound by an insulating cell membrane with a concentration of charged ions on either side that determines a capacitance Cm. Finally, a neuron responds to such a signal with a change in voltage, or an electrical potential energy difference between the cell and its surroundings, which is observed to sometimes result in a voltage spike called an action potential. This quantity, then, is the quantity of interest and is given by Vm.
Integrate-and-fire
One of the earliest models of a neuron was first investigated in 1907 by Louis Lapicque.[1] A neuron is represented in time by

which is just the time derivative of the law of capacitance, Q = CV. When an input current is applied, the membrane voltage increases with time until it reaches a constant threshold Vth, at which point a delta function spike occurs and the voltage is reset to its resting potential, after which the model continues to run. The firing frequency of the model thus increases linearly without bound as input current increases.
The model can be made more accurate by introducing a refractory period tref that limits the firing frequency of a neuron by preventing it from firing during that period. Through some calculus involving a Fourier transform, the firing frequency as a function of a constant input current thus looks like
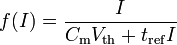 .
.
A remaining shortcoming of this model is that it implements no time-dependent memory. If the model receives a below-threshold signal at some time, it will retain that voltage boost forever until it fires again. This characteristic is clearly not in line with observed neuronal behavior.
Leaky integrate-and-fire
In the leaky integrate-and-fire model, the memory problem is solved by adding a "leak" term to the membrane potential, reflecting the diffusion of ions that occurs through the membrane when some equilibrium is not reached in the cell. The model looks like
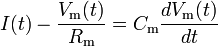
where Rm is the membrane resistance, as we find it is not a perfect insulator as assumed previously. This forces the input current to exceed some threshold Ith = Vth / Rm in order to cause the cell to fire, else it will simply leak out any change in potential. The firing frequency thus looks like
![f(I)={\begin{cases}0,&I\leq I_{{\mathrm {th}}}\\{[}t_{{\mathrm {ref}}}-R_{{\mathrm {m}}}C_{{\mathrm {m}}}\log(1-{\tfrac {V_{{\mathrm {th}}}}{IR_{{\mathrm {m}}}}}){]}^{{-1}},&I>I_{{\mathrm {th}}}\end{cases}}](/2014-wikipedia_en_all_02_2014/I/media/d/8/0/c/d80caa3a970d3e0efaf3d33f45bedf10.png)
which converges for large input currents to the previous leak-free model with refractory period.[2]
Exponential integrate-and-fire
In the Exponential Integrate-and-Fire, spike generation is exponential, following the equation:
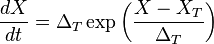 .
.
where  is the membrane potential,
is the membrane potential,  is the membrane potential threshold, and
is the membrane potential threshold, and 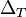 is the sharpness of action potential initiation, usually around 1 mV for cortical pyramidal neurons. Once the membrane potential crosses , it diverges to infinity in finite time.[3]
is the sharpness of action potential initiation, usually around 1 mV for cortical pyramidal neurons. Once the membrane potential crosses , it diverges to infinity in finite time.[3]
Hodgkin-Huxley
The most successful and widely used models of neurons have been based on the Markov kinetic model developed from Hodgkin and Huxley's 1952 work based on data from the squid giant axon. We note as before our voltage-current relationship, this time generalized to include multiple voltage-dependent currents:
 .
.
Each current is given by Ohm's Law as

where g(t,V) is the conductance, or inverse resistance, which can be expanded in terms of its constant average ḡ and the activation and inactivation fractions m and h, respectively, that determine how many ions can flow through available membrane channels. This expansion is given by
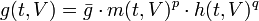
and our fractions follow the first-order kinetics

with similar dynamics for h, where we can use either τ and m∞ or α and β to define our gate fractions.
With such a form, all that remains is to individually investigate each current one wants to include. Typically, these include inward Ca2+ and Na+ input currents and several varieties of K+ outward currents, including a "leak" current. The end result can be at the small end 20 parameters which one must estimate or measure for an accurate model, and for complex systems of neurons not easily tractable by computer. Careful simplifications of the Hodgkin-Huxley model are therefore needed.
FitzHugh-Nagumo
Sweeping simplifications to Hodgkin-Huxley were introduced by FitzHugh and Nagumo in 1961 and 1962. Seeking to describe "regenerative self-excitation" by a nonlinear positive-feedback membrane voltage and recovery by a linear negative-feedback gate voltage, they developed the model described by
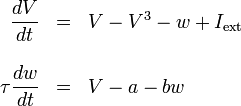
where we again have a membrane-like voltage and input current with a slower general gate voltage w and experimentally-determined parameters a = -0.7, b = 0.8, τ = 1/0.08. Although not clearly derivable from biology, the model allows for a simplified, immediately available dynamic, without being a trivial simplification.[4]
Morris-Lecar
In 1981 Morris and Lecar combined Hodgkin-Huxley and FitzHugh-Nagumo into a voltage-gated calcium channel model with a delayed-rectifier potassium channel, represented by
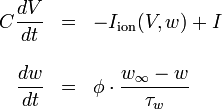
where 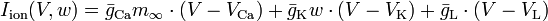 .[2]
.[2]
Hindmarsh-Rose
Building upon the FitzHugh-Nagumo model, Hindmarsh and Rose proposed in 1984 a model of neuronal activity described by three coupled first order differential equations:
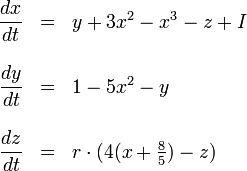
with r2 = x2 + y2 + z2, and r ≈ 10-2 so that the z variable only changes very slowly. This extra mathematical complexity allows a great variety of dynamic behaviors for the membrane potential, described by the x variable of the model, which include chaotic dynamics. This makes the Hindmarsh-Rose neuron model very useful, because being still simple, allows a good qualitative description of the many different patterns of the action potential observed in experiments.
Expanded neuron models
While the success of integrating and kinetic models is undisputed, much has to be determined experimentally before accurate predictions can be made. The theory of neuron integration and firing (response to inputs) is therefore expanded by accounting for the nonideal conditions of cell structure.
Cable theory
Cable theory describes the dendritic arbor as a cylindrical structure undergoing a regular pattern of bifurcation, like branches in a tree. For a single cylinder or an entire tree, the input conductance at the base (where the tree meets the cell body, or any such boundary) is defined as
 ,
,
where L is the electrotonic length of the cylinder which depends on its length, diameter, and resistance. A simple recursive algorithm scales linearly with the number of branches and can be used to calculate the effective conductance of the tree. This is given by
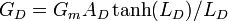
where AD = πld is the total surface area of the tree of total length l, and LD is its total electrotonic length. For an entire neuron in which the cell body conductance is GS and the membrane conductance per unit area is Gmd = Gm / A, we find the total neuron conductance GN for n dendrite trees by adding up all tree and soma conductances, given by
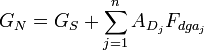 ,
,
where we can find the general correction factor Fdga experimentally by noting GD = GmdADFdga.
Compartmental models
The cable model makes a number of simplifications to give closed analytic results, namely that the dendritic arbor must branch in diminishing pairs in a fixed pattern. A compartmental model allows for any desired tree topology with arbitrary branches and lengths, but makes simplifications in the interactions between branches to compensate. Thus, the two models give complementary results, neither of which is necessarily more accurate.
Each individual piece, or compartment, of a dendrite is modeled by a straight cylinder of arbitrary length l and diameter d which connects with fixed resistance to any number of branching cylinders. We define the conductance ratio of the ith cylinder as Bi = Gi / G∞, where 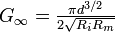 and Ri is the resistance between the current compartment and the next. We obtain a series of equations for conductance ratios in and out of a compartment by making corrections to the normal dynamic Bout,i = Bin,i+1, as
and Ri is the resistance between the current compartment and the next. We obtain a series of equations for conductance ratios in and out of a compartment by making corrections to the normal dynamic Bout,i = Bin,i+1, as
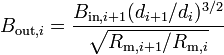
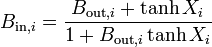
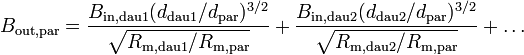
where the last equation deals with parents and daughters at branches, and 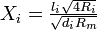 . We can iterate these equations through the tree until we get the point where the dendrites connect to the cell body (soma), where the conductance ratio is Bin,stem. Then our total neuron conductance is given by
. We can iterate these equations through the tree until we get the point where the dendrites connect to the cell body (soma), where the conductance ratio is Bin,stem. Then our total neuron conductance is given by
 .
.
Synaptic transmission
The response of a neuron to individual neurotransmitters can be modeled as an extension of the classical Hodgkin-Huxley model with both standard and nonstandard kinetic currents. Four neurotransmitters primarily have influence in the CNS. AMPA/kainate receptors are fast excitatory mediators while NMDA receptors mediate considerably slower currents. Fast inhibitory currents go through GABAA receptors, while GABAB receptors mediate by secondary G-protein-activated potassium channels. This range of mediation produces the following current dynamics:
![I_{{\mathrm {AMPA}}}(t,V)={\bar {g}}_{{\mathrm {AMPA}}}\cdot [O]\cdot (V(t)-E_{{\mathrm {AMPA}}})](/2014-wikipedia_en_all_02_2014/I/media/9/7/d/f/97dff92ca5198520700c40f5b0c02755.png)
![I_{{\mathrm {NMDA}}}(t,V)={\bar {g}}_{{\mathrm {NMDA}}}\cdot B(V)\cdot [O]\cdot (V(t)-E_{{\mathrm {NMDA}}})](/2014-wikipedia_en_all_02_2014/I/media/a/b/a/3/aba3a2d96878d87b272c20f7724b4066.png)
![I_{{\mathrm {GABA_{A}}}}(t,V)={\bar {g}}_{{\mathrm {GABA_{A}}}}\cdot ([O_{1}]+[O_{2}])\cdot (V(t)-E_{{\mathrm {Cl}}})](/2014-wikipedia_en_all_02_2014/I/media/e/c/7/5/ec7587ff04b85f97f9b5aa2f2235c7af.png)
![I_{{\mathrm {GABA_{B}}}}(t,V)={\bar {g}}_{{\mathrm {GABA_{B}}}}\cdot {\tfrac {[G]^{n}}{[G]^{n}+K_{{\mathrm {d}}}}}\cdot (V(t)-E_{{\mathrm {K}}})](/2014-wikipedia_en_all_02_2014/I/media/a/9/1/1/a911f3a6e209120de7979b26ae65d01f.png)
where ḡ is the maximal[5][2] conductance (around 1S) and E is the equilibrium potential of the given ion or transmitter (AMDA, NMDA, Cl, or K), while [O] describes the fraction of receptors that are open. For NMDA, there is a significant effect of magnesium block that depends sigmoidally on the concentration of intracellular magnesium by B(V). For GABAB, [G] is the concentration of the G-protein, and Kd describes the dissociation of G in binding to the potassium gates.
The dynamics of this more complicated model have been well-studied experimentally and produce important results in terms of very quick synaptic potentiation and depression, that is, fast, short-term learning.
Other conditions
The models above are still idealizations. Corrections must be made for the increased membrane surface area given by numerous dendritic spines, temperatures significantly hotter than room-temperature experimental data, and nonuniformity in the cell's internal structure.[2] Certain observed effects do not fit into some of these models. For instance, the temperature cycling (with minimal net temperature increase) of the cell membrane during action potential propagation not compatible with models which rely on modeling the membrane as a resistance which must dissipate energy when current flows through it. The transient thickening of the cell membrane during action potential propagation is also not predicted by these models, nor is the changing capacitance and voltage spike that results from this thickening incorporated into these models. The action of some anesthetics such as inert gases is problematic for these models as well. New models, such as the soliton model attempt to explain these phenomena, but are less developed than older models and have yet to be widely applied.
References
- ↑ Abbott, L.F. (1999). "Lapique's introduction of the integrate-and-fire model neuron (1907)". Brain Research Bulletin 50 (5/6): 303–304. doi:10.1016/S0361-9230(99)00161-6. PMID 10643408. Retrieved 2007-11-24.
- ↑ 2.0 2.1 2.2 2.3 Koch, Christof; Segev, Idan (1999). Methods in neuronal modeling : from ions to networks (2nd ed.). Cambridge, Mass.: MIT Press. p. 687. ISBN 0-262-11231-0.
- ↑ Ostojic, S.; Brunel, N.; Hakim, V. (2009). "How Connectivity, Background Activity, and Synaptic Properties Shape the Cross-Correlation between Spike Trains". Journal of Neuroscience 29 (33): 10234–10253. doi:10.1523/JNEUROSCI.1275-09.2009. PMID 19692598.
- ↑ Fitzhugh, R.; Izhikevich, E. (2006). "FitzHugh-Nagumo model". Scholarpedia 1 (9): 1349. doi:10.4249/scholarpedia.1349.
- ↑ Hodgkin, A. L.; Huxley, A. F. (1952). "A quantitative description of membrane current and its application to conduction and excitation in nerve". The Journal of physiology 117 (4): 500–544. PMC 1392413. PMID 12991237.