| Sea Urchin Temporal range: Ordovician–Recent |
|
|---|---|
| The sea urchin (Echinus melo) from Sardinia | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Echinodermata |
| Subphylum: | Echinozoa |
| Class: | Echinoidea Leske, 1778 |
| Subclasses | |
|
|
Sea urchins or urchins are small, spiny, globular animals which, with their close kin, such as sand dollars, constitute the class Echinoidea of the echinoderm phylum. They inhabit all oceans. Their shell, or "test", is round and spiny, typically from 3 to 10 centimetres (1.2 to 3.9 in) across. Common colors include black and dull shades of green, olive, brown, purple, and red. They move slowly, feeding mostly on algae. Sea otters, wolf eels, triggerfish, and other predators feed on them. Their "roe" (actually the gonads) is a delicacy in many cuisines.
The name "urchin" is an old name for the round spiny hedgehogs that sea urchins resemble.
Contents |
Sea urchins are members of the phylum Echinodermata, which also includes sea stars, sea cucumbers, brittle stars, and crinoids. Like other echinoderms they have fivefold symmetry (called pentamerism) and move by means of hundreds of tiny, transparent, adhesive "tube feet". The symmetry is not obvious in the living animal, but is easily visible in the dried test. "Echinodermate" means "spiny skin" in Greek.
Specifically, the term "sea urchin" refers to the "regular echinoids," which are symmetrical and globular. The term includes several different taxonomic groups: the order Echinoida, the order Cidaroida or "slate-pencil urchins", which have very thick, blunt spines, and others. Besides sea urchins, the class Echinoidea also includes three groups of "irregular" echinoids: flattened sand dollars, sea biscuits, and heart urchins.
Together with sea cucumbers (Holothuroidea), they make up the subphylum Echinozoa, which is characterized by a globoid shape without arms or projecting rays. Sea cucumbers and the irregular echinoids have secondarily evolved diverse shapes. Although many sea cucumbers have branched tentacles surrounding the oral opening, these have originated from modified tube feet and are not homologous to the arms of the crinoids, sea stars, and brittle stars.
Urchins typically range in size from 6 to 12 centimetres (2.4 to 4.7 in), although the largest species can reach up to 36 centimetres (14 in).[1]
Like other echinoderms, sea urchins are bilaterans. Their early larvae have bilateral symmetry[2] but they develop fivefold symmetry as they mature. This is most apparent in the "regular" sea urchins, which have roughly spherical bodies, with five equally-sized parts radiating out from the central axis. Several sea urchins, however, including the sand dollars, are oval in shape, with distinct front and rear ends, giving them a degree of bilateral symmetry. In these urchins, the upper surface of the body is slightly domed, but the underside is flat, while the sides are devoid of tube feet. This "irregular" body form has evolved to allow the animals to burrow through sand or other soft material.[1]
The lower half of a sea urchin's body is referred to as the oral surface, because it contains the mouth, while the upper half is the aboral surface. The internal organs are enclosed in a hard test composed of fused plates of calcium carbonate covered by a thin dermis and epidermis. The test is rigid, and divides into five ambulacral areas separated by five inter-ambulacral areas. Each of these areas consists of two rows of plates, so that the test includes twenty rows in total. The plates are covered in rounded tubercles, to which the spines are attached. The inner surface of the test is lined by peritoneum.[1]
Urchins have tube feet, which arise from the five ambulacral areas. ( The tube feet are moved by the water vascular system.)
The mouth lies in the centre of the oral surface in regular urchins, or towards one end of irregular urchins. It is surrounded by lips of softer tissue, with numerous small bony pieces embedded in it. This area, called the peristome, also includes five pairs of modified tube feet and, in many species, five pairs of gills. On the upper surface, opposite the mouth, is a region termed the periproct, which surrounds the anus. The periproct contains a variable number of hard plates, depending on species, one of which contains the madreporite.[1]
The sea urchin builds its spicules, the sharp crystalline "bones" that constitute the animal’s endoskeleton, in the larval stage. The fully formed spicule is composed of a single crystal with an unusual morphology. It has no facets and within 48 hours of fertilization assumes a shape that looks very much like the Mercedes-Benz logo.[3]
In other echinoderms, the endoskeleton is associated with a layer of muscle that allows the animal to move its arms or other body parts. This is entirely absent in sea urchins, which are unable to move in this way.
The spines, long and sharp in some species, protect the urchin from predators. The spines inflict a painful wound when they penetrate human skin, but are not dangerous. It is not clear if the spines are venomous (unlike the pedicellariae between the spines, which are venomous).[4]
Typical sea urchins have spines that are 1 to 3 centimetres (0.39 to 1.2 in) in length, 1 to 2 millimetres (0.039 to 0.079 in) thick, and not terribly sharp. Diadema antillarum, familiar in the Caribbean, has thin, potentially dangerous spines that can reach 10 to 30 centimetres (3.9 to 12 in) long.
Sea urchins are dioecious, having separate male and female sexes, although there is generally no easy way to distinguish the two, except for their location on the sea bottom. Males generally choose an elevated and exposed location so that their milt can be broadcast by sea currents. Females generally choose a low-lying location in sea bottom crevices, presumably so that the tiny larvae can have better protection from predators. Indeed, very small sea urchins are always found hiding underneath rocks. Regular sea urchins have five gonads, lying underneath the interambulacral regions of the test, while the irregular forms have only four, with the hindmost gonad being absent. Each gonad has a single duct, rising from the upper pole to open at a gonopore lying in one of the genital plates surrounding the anus. The gonads are lined with muscles underneath the peritoneum, and these allow the animal to squeeze its gametes through the duct and into the surrounding sea water, where fertilization takes place.[1]
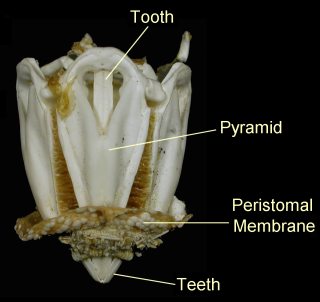
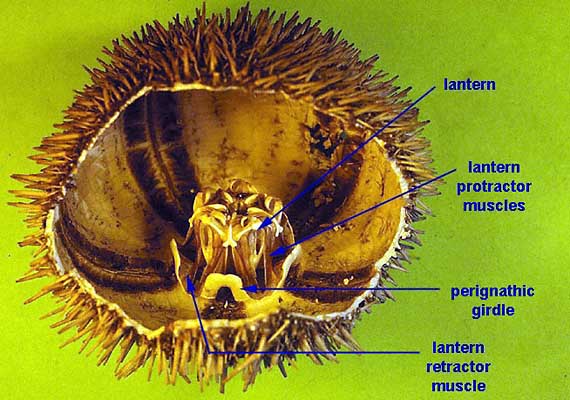
The mouth of most sea urchins is made up of five calcium carbonate teeth or jaws, with a fleshy tongue-like structure within. The entire chewing organ is known as Aristotle's lantern (image), from Aristotle's description in his History of Animals:
Recent research has shown that the sea urchin's teeth are self-sharpening; it can chew through stone.[5]
Heart urchins are unusual in not having a lantern. Instead, the mouth is surrounded by cilia that pull strings of mucus containing food particles towards a series of grooves around the mouth.[1]
The lantern, where present, surrounds both the mouth cavity and the pharynx. At the top of the lantern, the pharynx opens into the esophagus, which runs back down the outside of the lantern, to join the small intestine and a single caecum. The small intestine runs in a full circle around the inside of the test, before joining the large intestine, which completes another circuit in the opposite direction. From the large intestine, a rectum ascends towards the anus. Despite the names, the small and large intestine of sea urchins are in no way homologous to the similarly named structures in vertebrates.[1]
Digestion occurs in the intestine, with the caecum producing further digestive enzymes. An additional tube, called the siphon, runs beside much of the intestine, opening into it at both ends. It may be involved in resorption of water from food.[1]
Sea urchins possess both a water vascular system and a hemal system, the latter containing blood. However, the main circulatory fluid fills the general body cavity, or coelom. This fluid contains phagocytic coelomocytes which move through the vascular and hemal systems. The coelomocytes are an essential part of blood clotting, but also collect waste products and actively remove them from the body through the gills and tube feet.[1]
Most sea urchins possess five pairs of external gills, located around the mouth. These are thin-walled projections of the body cavity, and are the main organs of respiration in those urchins that possess them. Fluid can be pumped through the gills' interior by muscles associated with the lantern, but this is not continuous, and occurs only when the animal is low on oxygen. Tube feet can also act as respiratory organs, and are the primary sites of gas exchange in heart urchins and sand dollars, both of which lack gills.[1]
The nervous system of sea urchins has a relatively simple layout. There is no true brain. The center is a large nerve ring encircling the mouth just inside the lantern. From the nerve ring, five nerves radiate underneath the radial canals of the water vascular system, and branch into numerous finer nerves to innervate the tube feet, spines, and pedicellariae.[1]
Sea urchins are sensitive to touch, light, and chemicals. Although they do not have eyes or eye spots, recent research suggests that their entire body might function as one compound eye.[6] They also have statocysts, called spheridia, that are located within the ambulacral plates and help the animal remain upright.[1]
During early development, the sea urchin embryo undergoes ten cycles of cell division resulting in a single epithelial layer enveloping a blastocoel.[7] The embryo must then begin gastrulation, a multipart process which involves the dramatic rearrangement and invagination of cells to produce the three germ layers.
The first step of gastrulation is the epithelial to mesenchymal transition and ingression of primary mesenchyme cells into the blastocoel.[7] Primary mesenchyme cells, or PMCs, are cells located in the vegetal plate that are specified to become mesoderm.[8] Prior to ingression, PMCs exhibit all the features of other epithelial cells that comprise the embryo. Cells of the epithelium are bound basally to a laminal matrix and apically to an extra-embryonic matrix.[8] The apical microvilli of these cells reach into the hyaline layer, a component of the extra-embryonic matrix.[9] Neighboring epithelial cells are also connected to each other through apical junctions,[10] protein complexes containing adhesion molecules such as cadherins linked to catenins.
As PMCs begin to undergo an epithelial to mesenchymal transition, the lamina which binds them dissolves to begin the mechanical release of the cells.[9] Expression of the membrane protein that binds laminin, integrin, also becomes irregular at the beginning of ingression.[11] The microvilli which secure PMCs to the hyaline layer shorten,[12] as the cells reduce their affinity for the extra-embryonic matrix.[13] These cells concurrently increase their affinity for other components of the basal matrix, such as fibronectin, in part driving the movement of cells inward.[13] The apical junctions which bind PMCs to their neighboring epithelial cells become disrupted during this transition, and are absent in cells that have fully ingressed into the blastocoel.[9] Because staining for cadherins and catenins in ingressing cells decreases and develops as intracellular accumulations, apical junctions are thought to be cleared by endocytosis during ingression.[14][15]
Once the PMCs disrupt all attachment to their former location, the cells themselves change their morphology by contracting their apical surface, apical constriction, and enlarging their basal surface; acquiring a “bottle cell” phenotype.[8] Cytoskeletal rearrangements mediate the shape changes of PMCs; and though the cytoskeleton assists in the mechanics of ingression, other mechanisms drive the process. Experimentally disrupting microtubule dynamics in the species Strongylocentrotus pupuratus by applying colchicine stalls the ingression of PMCs but does not inhibit it.[16] Similarly, experimentally disrupting actin-myosin contraction using inhibitors slows down ingression, but does not arrest the process.[17]
The morphogenetic movements of the PMCs are an autonomous cellular behavior. Experimentally grafting PMCs into heterotopic tissue does not prevent the cells from ingressing.[13] In studies where PMCs are cultured in insolation, the cells were observed to gain affinity for fibronectin and simultaneously lose affinity for extra-embryonic matrix independent of the embryonic environment.[13]
At first glance, sea urchins often appear sessile, i.e. incapable of moving. Sometimes the most visible life sign is the spines, which attach to ball-and-socket joints and can point in any direction. In most urchins, touch elicits a prompt reaction from the spines, which converge toward the touch point. Sea urchins have no visible eyes, legs, or means of propulsion, but can move freely over hard surfaces using adhesive tube feet, working in conjunction with the spines.
In most cases, the eggs float freely in the sea, but some species hold onto them with their spines, affording them a greater degree of protection. The fertilized egg develops into a free-swimming blastula embryo in as little as twelve hours. Initially a simple ball of cells, the blastula soon transforms into a cone-shaped echinopluteus larva. In most species, this larva has twelve elongated arms, but in a few it contains supplies of nutrient yolk and lacks arms, since it has no need to feed. The arms are lined with bands of cilia that capture food particles and transport them to the mouth.[1]
It may take several months for the larva to complete its development, which begins with the formation of the test plates around the mouth and anus. Soon the larva sinks to the bottom and metamorphoses into adult form in as little as one hour. In some species, adults reach their maximum size in about five years.[1]
Sea urchins feed mainly on algae, but can also feed on sea cucumbers, and a wide range of invertebrates such as mussels, polychaetes, sponges, brittle stars and crinoids.[18] Population density varies by habitat with more dense populations being found in barren areas as compared to kelp stands.[19][20] Even in these barren areas, greatest densities are also found in shallow water. Populations are also generally found in deeper water if waves action is present.[20] Density also decrease in winter when storms cause them to seek protection in cracks and around larger underwater structures.[20] The shingle urchin (Colobocentrotus atratus) which lives on exposed shorelines is particularly resistant to wave action.[21]
Sea urchin is one of the favorite foods of sea otters and is also the main source of nutrition for wolf eels. Left unchecked, urchins devastate their environment, creating what biologists call an urchin barren, devoid of macroalgae and associated fauna. Sea otters have re-entered British Columbia, dramatically improving coastal ecosystem health.[22]
The earliest echinoid fossils date to the upper part of the Ordovician period (c 450 MYA), and the taxon has survived to the present day as a successful and diverse group of organisms. Spines may be present in well-preserved specimens, but usually only the test remains. Isolated spines are common as fossils. Some echinoids (such as Tylocidaris clavigera, from the Cretaceous period's English Chalk Formation) had very heavy club-shaped spines that would be difficult for an attacking predator to break through and make the echinoid awkward to handle. Such spines simplify walking on the soft sea-floor.
Most of the fossil echinoids from the Paleozoic era are incomplete, consisting of isolated spines and small clusters of scattered plates from crushed individuals. Most specimens occur in Devonian and Carboniferous rocks. The shallow water limestones from the Ordovician and Silurian periods of Estonia are famous for echinoids. Paleozoic echinoids probably inhabited relatively quiet waters. Because of their thin test, they would certainly not have survived in the wave-battered coastal waters inhabited by many modern echinoids. During the upper part of the Carboniferous period, there was a marked decline in echinoid diversity, and this trend continued to the Permian period. They neared extinction at the end of the Paleozoic era, with just six species known from the Permian period. Only two lineages survived this period's massive extinction of and into the Triassic: the genus Miocidaris, which gave rise to modern cidaroida (pencil urchins), and the ancestor that gave rise to the euechinoids. By the upper part of the Triassic period, their numbers began to increase again. Cidaroids have changed very little since the Late Triassic and are today considered to be living fossils.
The euechinoids, on the other hand, diversified into new lineages throughout the Jurassic period and into the Cretaceous period, and from them emerged the first irregular echinoids (superorder Atelostomata) during the early Jurassic, and later the other superorder (Gnathostomata) of irregular urchins which evolved independently. These two superorders are today representing 47% of all extant species of echinoids thanks to their adaptive breakthroughs, which allowed them to exploit habitats and food sources unavailable to regular echinoids. During the Mesozoic and Cenozoic eras the echinoids flourished. Most echinoid fossils are often abundant in the restricted localities and formations where they occur. An example of this is Enallaster, which exists by the thousands in certain outcrops of limestone from the Cretaceous period in Texas. Many fossils of the Late Jurassic Plesiocidaris still have the spines attached.
Some echinoids, such as Micraster which is found in the Cretaceous period Chalk Formation of England and France, serve as zone or index fossils. Because they evolved rapidly, they aid geologists in dating the surrounding rocks. However, most echinoids are not abundant enough and are of too limited range to serve as zone fossils.
In the early Tertiary (c 65 to 1.8 MYA), sand dollars (order Clypeasteroida) arose. Their distinctive flattened test and tiny spines were adapted to life on or under loose sand. They form the newest branch on the echinoid tree.
Sea urchins are traditional model organisms in developmental biology. This use originates from the 1800s, when their embryonic development became easily viewed by microscopy. Sea urchins were the first species in which sperm cells were proven to fertilize the ovum.
The recent sequencing of the sea urchin genome establishes homology between sea urchin and vertebrate immune system-related genes. Sea urchins code for at least 222 Toll-like receptor (TLR) genes and over 200 genes related to the vertebrates' Nod-like-receptor (NLR) family found in vertebrates.[23] This increases its usefulness as a valuable model organism for studying the evolution of innate immunity.
The gonads of both the male and female sea urchin, usually called sea urchin roe or corals,[24] are culinary delicacies in many parts of the world.[25][26]
In cuisines around the Mediterranean, Paracentrotus lividus is often eaten raw, with lemon.[27] It can also flavor omelettes, scrambled eggs, fish soup,[28] mayonnaise, Béchamel sauce for tartlets,[29] the boullie for a soufflé,[30] or Hollandaise sauce to make a fish sauce.[31] In Chile, it is served raw with lemon, onions, and olive oil.
Though the edible Strongylocentrotus droebachiensis is found in the North Atlantic, it is not widely eaten, though it is exported, mostly to Japan;[32] in Maine, sea urchins are known as whores' eggs. It was formerly a delicacy in the Orkney Islands, used instead of butter.[33]
In the West Indies, Cidaris tribuloides is eaten.[33]
On the Pacific Coast of North America, Strongylocentrotus franciscanus was praised by Euell Gibbons; Strongylocentrotus purpuratus is also eaten.
In New Zealand, Evechinus chloroticus, known as kina in Maori, is a delicacy, traditionally eaten raw. Though New Zealand fishermen would like to export them to Japan, their quality is too variable.[34]
In Japan, sea urchin is known as uni (ウニ), and its roe can retail for as much as $450/kg.;[35] it is served raw as sashimi or in sushi, with soy sauce and wasabi. Japan imports large quantities from the United States, South Korea, and other producers. Japanese demand for sea urchin corals has raised concerns about overfishing.[36]
|
|||||||||||||||||||||||||||||||||||||
{kind=link}
{kind=link}