Serpin

Serpins are a group of proteins with similar structures that were first identified as a set of proteins able to inhibit proteases. The acronym serpin was originally coined because many serpins inhibit chymotrypsin-like serine proteases (serine protease inhibitors).[2][3][4]
The first members of the serpin superfamily to be extensively studied were the human plasma proteins antithrombin and antitrypsin, which play key roles in controlling blood coagulation (e.g. Figure 1) and inflammation, respectively. Initially, research focused upon their role in human disease: antithrombin deficiency results in thrombosis and antitrypsin deficiency causes emphysema. In 1980 Hunt and Dayhoff made the surprising discovery that both these molecules share significant amino acid sequence similarity to the major protein in chicken egg white, ovalbumin, and they proposed a new protein superfamily.[5] Over 1000 serpins have now been identified, these include 36 human proteins, as well as molecules in plants, fungi, bacteria, archaea and certain viruses.[6][7][8] Serpins are thus the largest and most diverse family of protease inhibitors.[9]
While most serpins control proteolytic cascades, certain serpins do not inhibit enzymes, but instead perform diverse functions such as storage (ovalbumin, in egg white), hormone carriage proteins (thyroxine-binding globulin, cortisol-binding globulin) and tumor suppressor genes (maspin). The term serpin is used to describe these latter members as well, despite their noninhibitory function.[10]
As serpins control processes such as coagulation and inflammation, these proteins are the target of medical research. However, serpins are also of particular interest to the structural biology and protein folding communities, because they undergo a unique and dramatic change in shape (or conformational change) when they inhibit target proteases.[11] This is unusual - most classical protease inhibitors function as simple "lock and key" molecules that bind to and block access to the protease active site (see, for example, bovine pancreatic trypsin inhibitor). While the serpin mechanism of protease inhibition confers certain advantages, it also has drawbacks, and serpins are vulnerable to mutations that result in protein misfolding and the formation of inactive long-chain polymers (serpinopathies).[12][13][14] Serpin polymerisation reduces the amount of active inhibitor, as well as accumulation of serpin polymers, causing cell death and organ failure. For example, the serpin antitrypsin is primarily produced in the liver, and antitrypsin polymerisation causes liver cirrhosis.[14] Understanding serpinopathies also provides insights on protein misfolding in general, a process common to many human diseases, such as Alzheimer’s and CJD.[13]
Cross-class inhibitors

Most inhibitory serpins target chymotrypsin-like serine proteases (see Table 1 and Figure 2). These enzymes are defined by the presence of a nucleophilic serine residue in their catalytic site. Examples include thrombin, trypsin, and human neutrophil elastase.[16]
Some serpins inhibit other classes of protease and are termed "cross-class inhibitors". A number of such serpins have been shown to target cysteine proteases. These enzymes differ from serine proteases in that they are defined by the presence of a nucleophilic cysteine residue, rather than a serine residue, in their catalytic site.[17] Nonetheless, the enzymatic chemistry is similar, and serpins most likely inhibit both classes of enzyme in a similar fashion.[18]
Examples of cross-class inhibitory serpins include squamous cell carcinoma antigen 1 (SCCA-1) and the avian serpin myeloid and erythroid nuclear termination stage-specific protein (MENT) both inhibit papain-like cysteine proteases[19][20][21]
The viral serpin crmA is a suppressor of the inflammatory response through inhibition of IL-1 and IL-18 processing by the cysteine protease caspase-1.[22] In eukaryotes, a plant serpin has been shown to inhibit metacaspases. It is presently unclear whether any mammalian serpins function to inhibit caspases in vivo.
Localisation and roles
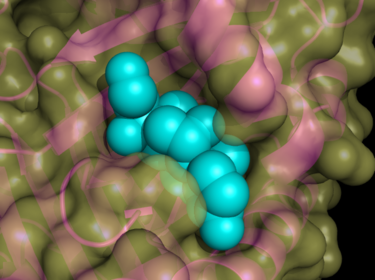
Approximately two-thirds of human serpins perform extracellular roles. For example, extracellular serpins regulate the proteolytic cascades central to blood clotting (antithrombin), the inflammatory response (antitrypsin, antichymotrypsin, and C1 inhibitor) and tissue remodeling (PAI-1). Non-inhibitory extracellular serpins also perform important roles. Thyroxine-binding globulin and cortisol-binding globulin transport the sterol hormones thyroxine and cortisol, respectively (Figure 3).[23][24] The protease renin cleaves off a ten-amino acid N-terminal peptide from angiotensinogen to produce the peptide hormone angiotensin I.[25] Table 1 provides a brief summary of human serpin function, as well as some of the diseases that result from serpin deficiency.
The first Intracellular members of the serpin superfamily were identified in the early 1990s.[26][27] As all nine serpins in Caenorhabditis elegans lack signal sequences, they are probably intracellular.[28] Based upon these data it seems likely that the ancestral serpin to human serpins was an intracellular molecule.
The protease targets of intracellular inhibitory serpins have been more difficult to identify. Characterization is complicated by the observation that many of these molecules appear to perform overlapping roles. Further many human serpins lack precise functional equivalents in model organisms such as the mouse. An important function of intracellular serpins may be to protect against the inappropriate activity of proteases inside the cell.[29] For example, one of the best-characterised human intracellular serpins is SERPINB9, which inhibits the cytotoxic granule protease granzyme B. In doing so, SERPINB9 may protect against inadvertent release of granzyme B and premature or unwanted activation of cell death pathways.[30]
Intracellular serpins also perform roles distinct from protease inhibition. For example, maspin, a non-inhibitory serpin, is important for preventing metastasis in breast and prostate cancers.[31][32] Another example is the avian nuclear cysteine protease inhibitor MENT, which acts as a chromatin remodelling molecule in avian red blood cells.[20][33]
Phylogenetic studies show that most intracellular serpins belong to a single clade (see Table 1). Exceptions include the non-inhibitory heat shock serpin HSP47, which is a chaperone essential for proper folding of collagen, and cycles between the cis-Golgi and the endoplasmic reticulum.[34]
Structure

Structural biology has played a central role in the understanding of serpin function and biology. Over eighty serpin structures, in a variety of different conformations (described below), have been determined to date. Although the function of serpins varies widely, these molecules all share a common structure (or fold).
The structure of the non-inhibitory serpin ovalbumin, and the inhibitory serpin antitrypsin, revealed the archetype native serpin fold.[37][38] All typically have three β-sheets (termed A, B and C) and eight or nine α-helices (hA-hI) (see figure 4). Serpins also possess an exposed region termed the reactive centre loop (RCL) that, in inhibitory molecules, includes the specificity determining region and forms the initial interaction with the target protease. In antitrypsin, the RCL is held at the top of the molecule and is not pre-inserted into the A β-sheet (figure 4, left panel). This conformation commonly exists in dynamic equilibrium with a partially inserted native conformation[39] seen in other inhibitory serpins (see figure 4, right panel).
Conformational change and inhibitory mechanism
Early studies on serpins revealed that the mechanism by which these molecules inhibit target proteases appeared distinct from the lock-and-key-type mechanism utilised by small protease inhibitors such as the Kunitz-type inhibitors (e.g. Basic pancreatic protease inhibitor). Indeed, serpins form covalent complexes with target proteases.[40] Structural studies on serpins also revealed that inhibitory members of the family undergo an unusual conformational change, termed the Stressed to Relaxed (S to R) transition.[37][39][41][42] During this structural transition the RCL inserts into β-sheet A (in red in figure 4 and 5) and forms an extra (fourth) β-strand. The serpin conformational change is key to the mechanism of inhibition of target proteases.
When attacking a substrate, serine proteases catalyze peptide bond cleavage in a two-step process. Initially, the catalytic serine performs a nucleophilic attack on the peptide bond of the substrate (Figure 5). This releases the new N-terminus and forms an ester-bond between the enzyme and the substrate. This covalent enzyme-substrate complex is called an acyl enzyme intermediate. Subsequent to this, this ester bond is hydrolysed and the new C-terminus is released. The RCL of a serpin acts as a substrate for its cognate protease. However, after the RCL is cleaved, but prior to hydrolysis of the acyl-enzyme intermediate, the serpin rapidly undergoes the S-to-R transition. Since the RCL is still covalently attached to the protease via the ester bond, the S-to-R transition causes the protease to be moved from the top to the bottom of the serpin. At the same time, the protease is distorted into a conformation, where the acyl enzyme intermediate is hydrolysed extremely slowly.[11] The protease thus remains covalently attached to the target protease and is thereby inhibited. Further, since the serpin has to be cleaved to inhibit the target protases, inhibition consumes the serpin as well. Serpins are therefore irreversible enzyme inhibitors. The serpin mechanism of inhibition is illustrated in figures 5 and 6, and several movies illustrating the serpin mechanism can be viewed at this link.

Figure 5:
Left: Structure of the non-covalent complex between insect Serpin1K and inactive rat trypsin (pdb code 1K9O).[43] To trap the encounter complex the trypsin (orange) was mutated to an inactive form unable to cleave the RCL. Serpin colouring is as for figure 4. Right: Final complex between antitrypsin and active trypsin (pdb code 1EZX).[11] The figure was produced using PYMOL. |

Figure 6: Catalytic mechanism of serine proteases (adapted from Serine protease mechanism) illustrating the stage in the cycle that is trapped by serpin inhibitors (magenta circle). The ester bond in the acyl enzyme intermediate is highlighted in red.
|
Conformational modulation of serpin activity
The conformational mobility of serpins provides a key advantage over static lock-and-key protease inhibitors. In particular, the function of inhibitory serpins can be readily controlled by specific cofactors. The X-ray crystal structures of antithrombin, heparin cofactor II, MENT and murine antichymotrypsin reveal that these serpins adopt a conformation wherein the first two amino acids of the RCL are inserted into the top of the A β-sheet (see figures 4 and 7). The partially-inserted conformation is important because co-factors are able to conformationally switch certain partially-inserted serpins into a fully-expelled form.[44][45] This conformational rearrangement makes the serpin a more effective inhibitor.
The archetypal example of this situation is antithrombin, which circulates in plasma in a partially-inserted relatively inactive state. The primary specificity determining residue (the P1 Arginine) points toward the body of the serpin and is unavailable to the protease (Figure 7). Upon binding a high-affinity heparin pentasaccharide sequence within long-chain heparin, antithrombin undergoes a conformational change, RCL expulsion, and exposure of the P1 Arginine. The heparin pentasaccharide-bound form of antithrombin is, thus, a more effective inhibitor of thrombin and factor Xa (figure 7).[46][47] Furthermore, both of these coagulation proteases contain binding sites (called exosites) for heparin. Heparin, therefore, also acts as a template for binding of both protease and serpin, further dramatically accelerating the interaction between the two parties (Figure 7). After the initial interaction, the final serpin complex is formed and the heparin moiety is released. This interaction is physiologically important. For example, after injury to the blood vessel wall, heparin is exposed, and antithrombin is activated to control the clotting response. The understanding of the molecular basis of this interaction formed the basis of the development of Fondaparinux, a synthetic form of Heparin pentasaccharide used as an anti-clotting drug.[48]

From left to right.
1. The partially inserted conformation of native antithrombin. The P1 Arginine is in purple spheres (from pdb 2ANT).
2. Binding of the high affinity heparin pentasaccharide sequence (in cyan spheres) within long chain heparin (in yellow spheres) (from pdb 1TB6).
Note how the P1 Arginine residue has flipped to a more exposed position.
3. Initial interaction of thrombin (orange) with the RCL. Thrombin also contains a binding site for heparin (from pdb 1TB6).
4. Following docking, the final serpin enzyme complex is formed (illustrated using the antitrypsin / trypsin complex) and heparin is released (from pdb 1EZX).
Certain serpins spontaneously undergo the S-to-R transition as part of their function, to form a conformation termed the latent state (Figure 8). In latent serpins, the first strand of the C-sheet has to peel off to allow full RCL insertion. Latent serpins are unable to interact with proteases and are not protease inhibitors. The transition to latency represents a control mechanism for the serpin PAI-1. PAI-1 is released in the inhibitory conformation, however, undergoes conformational change to the latent state unless it is bound to the cofactor vitronectin.[49] Thus PAI-1 contains an "auto-inactivation" mechanism. Similarly, antithrombin can also spontaneously convert to the latent state as part of its normal function. Finally, the N-terminus of tengpin (see pdbs 2PEE and 2PEF), a serpin from Thermoanaerobacter tengcongensis, is required to lock the molecule in the native inhibitory state. Disruption of interactions made by the N-terminal region results in spontaneous conformational change of this serpin to the latent conformation.[50][51]

Figure 8b: Structure of native PAI-1 bound to vitronectin (in cyan) (from pdb 1OCO). Part of the RCL is disordered in this structure and is represented by a dashed line.
Serpin receptor interactions
In humans, extracellular serpin-enzyme complexes are rapidly cleared from circulation. In mammals, one mechanism by which this occurs is via the low-density lipoprotein receptor-related protein (LRP receptor), which binds to inhibitory complexes made by antithrombin, PA1-1, and neuroserpin, causing uptake and subsequent signaling events.[52][53] Thus, as a consequence of the conformational change during serpin-enzyme complex formation, serpins may act as signaling molecules that alert cells to the presence of protease activity.[52] The fate of intracellular serpin-enzyme complexes remains to be characterised.
Recently, it has been shown that the Drosophila serpin necrotic is taken up via the Lipophorin Receptor-1 (LpR1), which is related to the mammalian LDL receptor family. Trafficking studies reveal that following uptake by LpR1, necrotic is delivered to lysosomes where it is targeted for degradation.[54]
Conformational change and non-inhibitory function
Certain non-inhibitory serpins also use the serpin conformational change as part of their function. For example, the native (S) form of thyroxine-binding globulin has high affinity for thyroxine, whereas the cleaved (R) form has low affinity. In similar manner, native (S) Cortisol-Binding Globulin (CBG) has higher affinity for cortisol than its cleaved (R) counterpart (Figure 3). Thus, in these serpins, RCL cleavage and the S to R transition has been commandeered to allow for ligand release, rather than protease inhibition.[23][24][55]
Serpins, serpinopathies and human disease

Serpins are vulnerable to inactivating disease-causing mutations that result in the formation of misfolded polymers or protein aggregates ("serpinopathies"). Well-characterised serpinopathies include alpha 1-antitrypsin deficiency (alpha-1), which may cause familial emphysema and sometimes liver cirrhosis, certain familial forms of thrombosis related to antithrombin deficiency, types 1 and 2 hereditary angioedema (HAE) related to deficiency of C1-inhibitor, and familial encephalopathy with neuroserpin inclusion bodies (FENIB; a rare type of dementia caused by neuroserpin polymerisation).[13][14] Serpins thus belong to a large group of molecules such as the prion proteins and the glutamine repeat containing proteins that cause proteopathies or conformational diseases.[13]
Serpin polymerisation causes disease in two ways. First, the lack of active serpin results in uncontrolled protease activity and tissue destruction; this is seen in the case of antitrypsin deficiency. Second, the polymers themselves clog up the endoplasmic reticulum of cells that synthesize serpins, eventually resulting in cell death and tissue damage. In the case of antitrypsin deficiency, antitrypsin polymers cause the death of liver cells, sometimes resulting in liver damage and cirrhosis. Within the cell, serpin polymers are removed via endoplasmic reticulum associated degradation.[57] However, the mechanism by which serpin polymers cause cell death remains to be fully understood.
Like cleaved serpins, serpin polymers are hyperstable with respect to heating, and each serpin monomer appears to have undergone the stressed to relaxed transition. Furthermore, serpin polymers are unable to inhibit target proteases, suggesting that the RCL is unavailable and inserted into the A-sheet. In the absence of definitive structural data, it was, therefore, postulated that serpins polymerise via a mechanism known as A-sheet polymerisation [14]. In normal function the RCL inserts into the A β-sheet to form a fourth strand (figure 4). In the A-sheet polymerisation model, it was suggested that the RCL of one serpin molecule spontaneously inserted into the A-sheet of another, to form a long-chain polymer (figure 9). In effect, it was, thus, proposed that polymerization occurred as a consequence of the requirement of the serpin scaffold to accept an additional β-strand.
Serpins were one of the first families for which disease-causing mutations were directly analyzed in reference to the available crystal structures.[58] In support of the A-sheet polymerisation model, it was noted that many serpin mutations that cause polymerisation localise to two distinct regions of the molecule (highlighted in figure 4a) termed the shutter and the breach. The shutter and the breach contain highly-conserved residues, underlie the path of RCL insertion, and are proposed to be important for conformational change.
Two structures of cleaved serpin polymers have been solved; both of which reveal RCL / A-sheet sheet linkages similar to those predicted by the A sheet polymerisation mechanism.[59][60] However, in direct contrast to the known properties of physiological serpin polymers, crystals of cleaved serpin A-sheet polymers readily dissociate into monomeric forms.[59][60]

A large body of data now suggest that the events associated with serpin polymerisation occur during the folding of the molecule, and that mutations that cause serpinopathies interfere with the ability of the serpin to fold to the metastable native state.[61] In normal serpin folding, the serpin rapidly moves through a key folding intermediate to attain the native state. Many studies have shown that it is the serpin folding intermediate that has the ability to polymerise, hence it is important that this folding species rapidly moves on to adopt native state. It was shown that mutations such as the Z-antitrypsin variant (Glu 342 to Lys) somehow prevented the final stage of seprin folding and caused the accumulation of the folding intermediate. As a result, population of the folding intermediate resulted in polymer formation.[61] Interestingly, it was noted that once folded, the Z-antitrypsin variant closely resembles wild-type material in terms of thermal stability and inhibitory activity.[61]
Together, these data have presented an important challenge to the A-sheet model for serpin polymerisation. On the one hand, the idea that serpin polymer formation essentially takes advantage of the serpin mechanism of conformational change is an attractive one. On the other, the biophyiscal data in particular suggest that it is a folding intermediate (rather than the native form) that polymerises, and it is clear that this intermediate must have different structural properties to the native, folded state.
In 2008, a key serpin crystal structure was determined that strongly suggests that physiological serpin polymers do not form via the A-sheet mechanism and instead form via a more extensive domain swapping event.[12] The structure solved is of an antithrombin dimer (figure 10), and reveals that both strands s5A and the RCL are able to be incorporated into the A-sheet of another serpin molecule. This structure can readily be adapted to form long chain polymers (figure 11).[12]
The new "domain swapped" model for serpin polymerisation reconciles the available biophysical and biochemical data. In particular, these data suggest that the final stage of serpin folding to the native state is most likely the incorporation of the fifth strand (s5A). Of key importance is the observation that several polymerogenic mutations (including the Z-variant) cluster on and around s5A and mutation of these residues may prevent proper incorporation of s5A into the A-sheet. As a result, during folding the mutation causes the serpin to remain "stuck" in the intermediate form. Much of the intermediate species, unable to efficiently form the native conformation, eventually forms hyperstable polymers via the insertion of both s5A and the RCL into another intermediate (figure 11).

Mutations that result in spontaneous formation of latent (or latent-like), inactive conformations

Certain pathogenic mutations in serpins can promote inappropriate transition to the monomeric latent state (see figure 8a for the structure of the latent state). This causes disease because it reduces the amount of active inhibitory serpin. For example, the disease-linked antithrombin variants wibble and wobble,[63] both promote formation of the latent state.
It is also worth highlighting a structure of a disease-linked human antichymotrypsin variant that further demonstrates the extraordinary flexibility of the serpin scaffold. The structure of antichymotrypsin (Leucine 55 to Proline) revealed a novel "δ" conformation that may represent an intermediate between the native and latent state (Figure 12). In the delta conformation, four residues of the RCL are inserted into the top of β-sheet A. The bottom half of the sheet is filled as a result of one of the α-helices (the F-helix) partially switching to a β-strand conformation, completing the β-sheet hydrogen bonding.[62] It is unclear whether other serpins can adopt this conformer, and whether this conformation has a functional role. However, it is speculated that the δ-conformation may be adopted by Thyroxine-binding globulin during thyroxine release.[24]
In humans, simple deficiency of many serpins (e.g., through a null mutation) may result in disease (see Table 1).
It is rare that single amino acid changes in the RCL of a serpin alters the specificity of the inhibitor and allow it to target the wrong protease. For example, the Antitrypsin-Pittsburgh mutation (methionine 358 to arginine) allowed the serpin to inhibit thrombin, thus causing a bleeding disorder.[64]
Serpins are suicide inhibitors, the RCL acting as a "bait." Certain disease-linked mutations in the RCL of human serpins permit true substrate-like behaviour and cleavage without complex formation. Such variants are speculated to affect the rate or the extent of RCL insertion into the A-sheet. These mutations, in effect, result in serpin deficiency through a failure to properly control the target protease.[58][65]
Several non-inhibitory serpins play key roles in important human diseases. For example, maspin functions as a tumour suppressor in breast and prostate cancer. The mechanism of maspin function remains to be fully understood. Murine knockouts of maspin are lethal; these data suggest that maspin plays a key role in development.[66]
Evolution
Serpins were initially believed to be restricted to eukaryote organisms, but have since been found in a number of bacteria and archaea.[6][7][67] It remains unclear whether these prokaryote genes are the descendants of an ancestral prokaryotic serpin or the product of lateral gene transfer (genetic transfer between organisms not by evolutionary descent). Rawlings et al. showed that serpins are the most widely-distributed and largest family of protease inhibitors.[9]
Types of serpin
Human serpins
In 2001, a serpin nomenclature was established (see table 1, below).[10] The naming system is based upon a phylogenetic analysis of ~500 serpins.[6] The human genome encodes 16 serpin clades, termed serpinA through to serpinP, encoding 29 inhibitory and 7 non-inhibitory serpin proteins (see Law et al. (2006) for a recent review).[68] The proteins are named serpinXY where X is the clade of the protein and Y the number of the protein within that clade. Table 1 lists each human serpin, together with brief notes in regards to each molecules function and the consequence (where known) of dysfunction or deficiency.
Table 1
| Protein name | PDB | Common Name | Description | Disease / Effect of deficiency | Chromosomal location |
| SERPINA1 | 1QLP 7API 1D5S |
Alpha 1-antitrypsin | extracellular, inhibits human neutrophil elastase.[69] | Deficiency results in emphysema, antitrypsin polymerisation results in cirrhosis. Serpinopathy.[14] The C-terminal fragment of cleaved SERPINA1 may inhibit HIV-1 infection.[70] | 14q32.1 |
| SERPINA2 | Antitrypsin-related protein | extracellular, possible pseudogene[71] | Unknown | 14q32.1 | |
| SERPINA3 | 1YXA 2ACH[72] |
Alpha 1-antichymotrypsin | Extracellular, inhibits cathepsin G.[73] | Deficiency results in emphysema. Serpinopathy[62] | 14q32.1 |
| SERPINA4 | Kallistatin | extracellular, inhibition of kallikrein, regulation of vascular function[74][75] | Unknown | 14q32.1 | |
| SERPINA5 | 2OL2[76] 3B9F[77] |
Protein C inhibitor | Extracellular, inhibitor of active protein C.[78] Intracellular role in preventing phagocytosis of bacteria.[79] | Male murine knockouts are infertile[80] In multiple sclerosis, accumulation of PCI has been noted in chronic active plaques.[81] | 14q32.1 |
| SERPINA6 | 2V6D[23] 2VDX 2VDY |
Cortisol binding globulin | Extracellular, non-inhibitory; cortisol binding.[23] | Deficiency may cause chronic fatigue[82] | 14q32.1 |
| SERPINA7 | 2CEO[24] 2RIV 2RIW |
Thyroxine-binding globulin | extracellular, non-inhibitory; thyroxine binding[24] | Deficiency causes hypothyroidism.[83] | Xq22.2 |
| SERPINA8 | 2X0B 2WXW 2WXX 2WXY 2WXZ 2WY0 2WY1 |
Angiotensinogen | Extracellular; non-inhibitory, cleavage by renin results in release of angiotensin I.[84] | Variants linked to hypertension[85] Murine knockouts result in hypotension.[86] | 1q42-q43 |
| SERPINA9 | Centerin | Extracellular; inhibitory, maintenance of naive B cells[87][88] | Unknown | 14q32.1 | |
| SERPINA10 | 3F1S[1] 3H5C[89] |
Protein Z-related protease inhibitor | extracellular, binds protein Z and inactivates factor Xa and factor XIa)[90] | Deficiency may cause venous thromboembolic disease[91] | 14q32.1 |
| SERPINA11 | - | probably extracellular, not characterised. | Unknown | 14q32.13 | |
| SERPINA12 | Vaspin | extracellular, insulin-sensitizing adipocytokine[92] | Unknown | 14q32.1 | |
| SERPINA13 | - | probably extracellular, not characterised | Unknown | 14q32 | |
| SERPINB1 | 1HLE[93] | Monocyte neutrophil elastase inhibitor | Intracellular, inhibition of neutrophil elastase[94] | Murine knockout results in neutrophil survival defect and immune deficiency[95] | 6p25 |
| SERPINB2 | 1BY7[96] | Plasminogen activator inhibitor-2 | Intracellular/extracellular. Inhibition of extracellular uPA. Intracellular function unclear, however, may protect against viral infection.[97] | Murine knockouts viable / no obvious phenotype[98] | 18q21.3 |
| SERPINB3 | 2ZV6[99] | Squamous cell carcinoma antigen-1 (SCCA-1) | Intracellular, inhibitor of papain-like cysteine proteases[19] | Unknown | 18q21.3 |
| SERPINB4 | Squamous cell carcinoma antigen-2 (SCCA-2) | Intracellular, inhibitor of cathepsin G and chymase[100] | Unknown | 18q21.3 | |
| SERPINB5 | 1WZ9[101] | Maspin | Obligate intracellular serpin[102], non inhibitory, tumour suppressor in breast and prostate cancer[31] | Murine knockouts lethal, important role in cancer metastasis[66] | 18q21.3 |
| SERPINB6 | PI-6 | intracellular, inhibition of cathepsin G[103] | Murine knockout reveals mild neutropenia[104]. In humans, a nonsense mutation in SERPINB6 results in moderate to severe hearing loss[105]. | 6p25 | |
| SERPINB7 | Megsin | intracellular, involved in megakaryocyte maturation[106] | Unknown | 18q21.3 | |
| SERPINB8 | PI-8 | intracellular; possible furin inhibitor[107] | Unknown | 18q21.3 | |
| SERPINB9 | PI-9 | intracellular, inhibitor of the cytotoxic granule protease granzyme B[108] | murine knockout reveals immune dysfunction[109] | 6p25 | |
| SERPINB10 | Bomapin | intracellular, unknown function[110] | Analysis of murine genomic material (C57/BL6; the common lab strain) reveals a stop codon in this gene (BC069938). In contrast, EST data suggests that full length bomapin is expressed in Czech II mice. These data suggest that loss of Bomapin function in mice does not result in an overt phenotype. | 18q21.3 | |
| SERPINB11 | intracellular, unknown function[111] | Murine Serpinb11 is an active inhibitor whereas the human orthalogue is inactive.[111] | 18q21.3 | ||
| SERPINB12 | Yukopin | intracellular, unknown function[112] | Unknown | 18q21.3 | |
| SERPINB13 | Hurpin/Headpin | intracellular, inhibitor of papain-like cysteine proteases[113] | Unknown | 18q21.3 | |
| SERPINC1 | 2ANT 2ZNH 1AZX 1TB6 2GD4 1T1F |
Antithrombin | Extracellular, inhibitor of coagulation, specifically factor X, factor IX and thrombin[114] | Deficiency results in thrombosis and other clotting disorders. Serpinopathy[115] | 1q23-q21 |
| SERPIND1 | 1JMJ 1JMO[116] |
Heparin cofactor II | extracellular, thrombin inhibitor[117] | Murine knockouts are lethal.[118] | 22q11 |
| SERPINE1 | 1DVN[119] 1OC0[120] |
Plasminogen activator inhibitor 1 | Extracellular; inhibitor of thrombin, uPA and TPa.[121] | Cardiovascular disease, tumour progression[122][123] | 7q21.3-q22 |
| SERPINE2 | Glia derived nexin / Protease nexin I | Extracellular, inhibition of uPA and tPA.[124] | Abnormal expression leads to human male infertility.[125] Knockout mice also develop epileptic phenotype.[126] | 2q33-q35 | |
| SERPINF1 | 1IMV[127] | Pigment epithelium derived factor | Extracellular, non-inhibitory, potent anti-angiogenic molecule.[128] PEDF has been reported to bind the glycosaminoglycan hyaluronan.[129] | Murine knockout studies reveal that SERPINF1 regulates the vasculature and mass of the pancreas and the prostate.[128] Further, SERPINF1 has been demonstrated to promote Notch–dependent renewal of adult periventricular neural stem cells.[130] | 17p13.3 |
| SERPINF2 | 2R9Y[131] | Alpha 2-antiplasmin | extracellular, plasmin inhibitor, inhibitor of fibrinolysis.[132] | Bleeding disorder[133] | 17pter-p12 |
| SERPING1 | 2OAY[134] | Complement 1-inhibitor | Extracellular, C1 esterase inhibitor.[134] | Angiodemia, serpinopathy.[135] Several polymorphisms in the SERPING1 gene are strongly associated with development of age-related macular degeneration and blindness.[136] | 11q11-q13.1 |
| SERPINH1 | 47 kDa Heat shock protein (HSP47) | intracellular, non inhibitory, molecular chaperone in collagen folding.[137] | Murine knockouts are lethal.[138] A missense mutation in human SERPINH1 results in severe osteogenesis imperfecta.[139] | 11p15 | |
| SERPINI1 | 1JJO 3FGQ[140] 3F5N 3F02[141] |
Neuroserpin | Extracellular, inhibitor of tPA, uPA and plasmin[142] | Mutated in dementia (FENIB). Serpinopathy[143] | 3q26 |
| SERPINI2 | Pancpin | Extracellular, unknown protease target.[144] | Studies on the Pequeño mouse line revealed that loss of SERPINI2 results in pancreatic insufficiency through pancreatic acinar cell loss.[145] In addition a possible role for SERPINI2 in inhibition of pancreatic cancer metastasis has been suggested.[144] | 3q26 |
Insect Serpins
The Drosophila melanogaster genome contains 29 serpin encoding genes. Amino acid sequence analysis has placed 14 of these serpins in serpin clade Q and 3 in serpin clade K with the remaining 12 serpins classified as orphan serpins not belonging to any clade.[146] The clade classification system is difficult to use for Drosophila serpins and instead a nomenclature system has been adopted that is based on the position of Drosphila serpin genes on the Drosophila chromosomes. 13 of the Drosophila serpins occur as isolated genes in the genome (including Serpin-27A, see below), with the remaining 16 organised into three gene clusters that occur at chromosome positions 28D (2 serpins), 42D (5 serpins), 43A (4 serpins), 77B (3 serpins) and 88E (2 serpins).[146][147][148]
Drosophila serpin-27A
Studies on Drosophila serpins reveal that Serpin-27A inhibits the Easter protease (the final protease in the Nudel, Gastrulation Defective, Snake and Easter proteolytic cascade) and thus controls dorsoventral patterning. Easter functions to cleave Spätzle (a chemokine-type ligand), which results in Toll mediated signaling. In addition to its central role in embryonic patterning, Toll signaling is also important for the innate immune response in insects. Accordingly, serpin-27A additionally functions to control the insect immune response.[149][150][151]
Worm Serpins
The genome of the nematode worm C. elegans contains nine serpins, however, only five of these molecules appear to function as protease inhibitors.[28] One of these serpins, SRP-6, has been shown to perform a protective function and guard against stress induced calpain-associated lysosomal disruption. Further SRP-6 functions to inhibit lysosomal cysteine proteases released after lysosomal rupture. Accordingly, worms lacking SRP-6 are sensitive to stress. Most notably, SRP-6 knockout worms die when placed in water (the hypo-osmotic stress lethal phenotype or Osl). Based on these data it is suggested that lysosomes play a general and controllable role in determining cell fate.[152]
Plant serpins
The presence of serpins in plants has long been recognised [153], indeed, an abundant barley grain serpin (barley Protein Z) is one of the major protein components in beer.
The MEROPS database identifies 18 serpin family members in the Arabidopsis thaliana genome, but only about eight of these are full-length serpin sequences. Plant serpins are potent inhibitors of mammalian chymotrypsin-like serine proteases in vitro, the most well-studied example being barley serpin Zx (BSZx), which is able to inhibit trypsin, chymotrypsin as well as several blood coagulation factors [154]. However, close relatives of chymotrypsin-like serine proteases are absent in plants. Interestingly, the RCL of several serpins from wheat grain and rye [155] contain poly-Q repeat sequences similar to those present in the prolamin storage proteins of the endosperm.[156] It has therefore been suggested that plant serpins may function to inhibit proteases from insects or microbes that cleave grain storage proteins. In support of this hypothesis, specific plant serpins have been identified in the phloem sap of pumpkin (CmPS-1)[157] and cucumber plants [158][159]. However, while an inverse correlation between up-regulation of CmPS-1 expression and aphid survival was observed, in vitro feeding experiments revealed that recombinant CmPS-1 did not appear to affect insect survival[157].
Accordingly, alternative roles and protease targets for plant serpins have been proposed. Notably, it has recently been shown that Arabidopsis Serpin1 (At1g47710; 3LE2) inhibits metacaspase-like proteases and may control cell death pathways[160]. Two other Arabidopsis serpins, AtSRP2 (At2g14540) and AtSRP3 (At1g64030), are involved in responses to DNA damage caused by plant exposure to methane methylsulfonate (MMS)[161].
Fungal serpins
A single fungal serpin has been characterized to date: celpin from Piromyces sp. strain E2. Piromyces is an anaerobic fungus found in the gut of ruminants and is important for digesting plant material. Celpin is predicted to be an inhibitory molecule and contains two N-terminal dockerin domains in addition to the serpin domain. Dockerins are commonly found in proteins that localise to the fungal cellulosome, a large extracellular mulitprotein complex that breaks down cellulose[8]. It is therefore suggested that celpin protects the cellulosome against plant proteases. Interestingly certain bacterial serpins also localize to the cellulosome [162].
Prokaryote serpins
Predicted serpin genes are sporadically distributed in prokaryotes. In vitro studies on some of these moelcules have revealed that they are able to inhibit proteases and it is suggested that they function as inhibitors in vivo. Interestingly, several prokaryote serpins are found in extremophiles. Accordingly, and in contrast to mammalian serpins, these molecule possess elevated resistance to heat denaturation.[163][164] The precise role of most bacterial serpins remains obscure, however, Clostridium thermocellum serpin localises to the cellulosome. It is suggested that the role of cellulosome-associated serpins may be to prevent unwanted protease activity against the cellulosome.[162]
Viral serpins
Serpins are also expressed by viruses as a way to evade the host's immune defense.[165] In particular, serpins expressed by pox viruses, including cow pox (vaccinia) and rabbit pox (myxoma), are of interest because of their potential use as novel therapeutics for immune and inflammatory disorders as well as transplant therapy.[166][167] A study on Serp1 reveals this molecule suppresses the Toll-mediated innate immune response and allows indefinite cardiac allograft survival in rats.[166][168] Studies on Crma and Serp2, reveal both are cross-class inhibitor and targets both serine (Granzyme B; albeit weakly) and cysteine proteases (Caspase 1 and Caspase 8).[169][170] In comparison to their mammalian counterparts, viral serpins contain significant deletions of elements of secondary structure. Specifically, structural studies on crmA reveals this molecule lacks the D-helix as well as significant portions of the A- and E-helices.[171]
See also
- Wikipedia:MeSH D12.776#MeSH D12.776.872 --- serpins
- Proteopathy
- Familial encephalopathy with neuroserpin inclusion bodies
References
- ↑ 1.0 1.1 Wei Z, Yan Y, Carrell RW, Zhou A (June 2009). "Crystal structure of protein Z-dependent inhibitor complex shows how protein Z functions as a cofactor in the membrane inhibition of factor X". Blood 114 (17): 3662–7. doi:10.1182/blood-2009-04-210021. PMID 19528533.
- ↑ R. Carrell and J. Travis. (1985). "α1-Antitrypsin and the serpins: Variation and countervariation". Trends Biochem. Sci. 10: 20–24. doi:10.1016/0968-0004(85)90011-8.
- ↑ Whisstock JC, Silverman GA, Bird PI, Bottomley SP, Kaiserman D, Luke CJ, Pak SC, Reichhart JM, Huntington JA (May 2010). "Serpins flex their muscle: structural insights into target peptidase recognition, polymerization and transport functions". J Biol Chem 285 (32): 24307–12. doi:10.1074/jbc.R110.141408. PMID 20498368.
- ↑ Silverman GA, Whisstock JC, Bottomley SP, Huntington JA, Kaiserman D, Luke CJ, Pak SC, Reichhart JM, Bird PI (May 2010). "Serpins flex their muscle: putting the clamps on proteolysis in diverse biological systems". J Biol Chem 285 (32): 24299–305. doi:10.1074/jbc.R110.112771. PMID 20498369.
- ↑ Hunt LT, Dayhoff MO (1980). "A surprising new protein superfamily containing ovalbumin, antithrombin-III, and α1-proteinase inhibitor". Biochem Biophys Res Commun. 95 (2): 864–71. doi:10.1016/0006-291X(80)90867-0. PMID 6968211.
- ↑ 6.0 6.1 6.2 Irving JA, Pike RN, Lesk AM, Whisstock. (2000). "Phylogeny of the Serpin Superfamily: Implications of Patterns of Amino Acid Conservation for Structure and Function". Genome Res. 10 (12): 1845–64. doi:10.1101/gr.GR-1478R. PMID 11116082.
- ↑ 7.0 7.1 Irving J, Steenbakkers P, Lesk A, Op den Camp H, Pike R, Whisstock J (2002). "Serpins in prokaryotes". Mol Biol Evol 19 (11): 1881–90. PMID 12411597.
- ↑ 8.0 8.1 Steenbakkers PJ, Irving JA, Harhangi HR, et al. (August 2008). "A serpin in the cellulosome of the anaerobic fungus Piromyces sp. strain E2". Mycol. Res. 112 (Pt 8): 999–1006. doi:10.1016/j.mycres.2008.01.021. PMID 18539447.
- ↑ 9.0 9.1 Rawlings ND, Tolle DP, Barrett AJ. (2004). "Evolutionary families of peptidase inhibitors". Biochem J. 378 (Pt 3): 705–16.. doi:10.1042/BJ20031825. PMID 14705960.
- ↑ 10.0 10.1 Silverman GA, Bird PI, Carrell RW, Church FC, Coughlin PB, Gettins PG, Irving JA, Lomas DA, Luke CJ, Moyer RW, Pemberton PA, Remold-O'Donnell E, Salvesen GS, Travis J, Whisstock JC. (2001). "'The serpins are an expanding superfamily of structurally similar but functionally diverse proteins. Evolution, mechanism of inhibition, novel functions, and a revised nomenclature". J Biol Chem 276 (36): 33293–6. doi:10.1074/jbc.R100016200. PMID 11435447.
- ↑ 11.0 11.1 11.2 Huntington J, Read R, Carrell R (2000). "Structure of a serpin-protease complex shows inhibition by deformation". Nature 407 (6806): 923–6. doi:10.1038/35038119. PMID 11057674.
- ↑ 12.0 12.1 12.2 12.3 12.4 Yamasaki M, Li W, Johnson DJ, Huntington JA (October 2008). "Crystal structure of a stable dimer reveals the molecular basis of serpin polymerization". Nature 455 (7217): 1255. doi:10.1038/nature07394. PMID 18923394.
- ↑ 13.0 13.1 13.2 13.3 Carrell RW, Lomas DA. (1997). "Conformational disease". Lancet. 350 (9071): 134–8. doi:10.1016/S0140-6736(97)02073-4. PMID 9228977.
- ↑ 14.0 14.1 14.2 14.3 14.4 14.5 Lomas DA, Evans DL, Finch JT & Carrell RW (1992). "The mechanism of Z alpha 1-antitrypsin accumulation in the liver". Nature 357 (6379): 605–607. doi:10.1038/357605a0. PMID 1608473.
- ↑ Tsukada H, Blow DM (1985). "Structure of alpha-chymotrypsin refined at 1.68 A resolution". J. Mol. Biol. 184 (4): 703–11. doi:10.1016/0022-2836(85)90314-6. PMID 4046030.
- ↑ Barrett AJ, Rawlings ND. (1995). "Families and clans of serine peptidases". Arch Biochem Biophys. 318 (2): 247–50. doi:10.1006/abbi.1995.1227. PMID 7733651.
- ↑ Barrett AJ, Rawlings ND. (2001). "Evolutionary lines of cysteine peptidases". Biol Chem 382 (5): 727–33.. doi:10.1515/BC.2001.088. PMID 11517925.
- ↑ Irving JA, Pike RN, Dai W, Bromme D, Worrall DM, Silverman GA, Coetzer TH, Dennison C, Bottomley SP, Whisstock JC. (2002). "Evidence that serpin architecture intrinsically supports papain-like cysteine protease inhibition: engineering alpha(1)-antitrypsin to inhibit cathepsin proteases". Biochemistry. 41 (15): 4998–5004. doi:10.1021/bi0159985. PMID 11939796.
- ↑ 19.0 19.1 Schick C, Brömme D, Bartuski A, Uemura Y, Schechter N, Silverman G (1998). "The reactive site loop of the serpin SCCA1 is essential for cysteine proteinase inhibition". Proc Natl Acad Sci USA 95 (23): 13465–70. doi:10.1073/pnas.95.23.13465. PMID 9811823.
- ↑ 20.0 20.1 McGowan S, Buckle A, Irving J, Ong P, Bashtannyk-Puhalovich T, Kan W, Henderson K, Bulynko Y, Popova E, Smith A, Bottomley S, Rossjohn J, Grigoryev S, Pike R, Whisstock J (2006). "X-ray crystal structure of MENT: evidence for functional loop-sheet polymers in chromatin condensation". EMBO J 25 (13): 3144–55. doi:10.1038/sj.emboj.7601201. PMID 16810322.
- ↑ Ong PC, McGowan S, Pearce MC, Irving JA, Kan WT, Grigoryev SA, Turk B, Silverman GA, Brix K, Bottomley SP, Whisstock JC, Pike RN (2007). "DNA accelerates the inhibition of human cathepsin V by serpins". Journal of Biological Chemistry 282 (51): 36980. doi:10.1074/jbc.M706991200. PMID 17923478.
- ↑ Ray C, Black R, Kronheim S, Greenstreet T, Sleath P, Salvesen G, Pickup D (1992). "Viral inhibition of inflammation: cowpox virus encodes an inhibitor of the interleukin-1 beta converting enzyme". Cell 69 (4): 597–604. doi:10.1016/0092-8674(92)90223-Y. PMID 1339309.
- ↑ 23.0 23.1 23.2 23.3 23.4 Klieber MA, Underhill C, Hammond GL, Muller YA. (2007). "Corticosteroid-binding globulin: structural basis for steroid transport and proteinase-triggered release". J Biol Chem. 282 (40): 29594. doi:10.1074/jbc.M705014200. PMID 17644521.
- ↑ 24.0 24.1 24.2 24.3 24.4 Zhou A, Wei Z, Read RJ, Carrell RW. (2006). "Structural mechanism for the carriage and release of thyroxine in the blood". Proc Natl Acad Sci U S A. 103 (=36): 13321–6.. doi:10.1073/pnas.0604080103. PMID 16938877.
- ↑ Campbell DJ. (2003). "The renin-angiotensin and the kallikrein-kinin systems". Int J Biochem Cell Biol. 35 (6): 784–91. doi:10.1016/S1357-2725(02)00262-5. PMID 12676165.
- ↑ Remold-O'Donnell E, Chin J, Alberts M. (1992). "Sequence and molecular characterization of human monocyte/neutrophil elastase inhibitor". Proc Natl Acad Sci U S A. 89 (12): 5635–9. doi:10.1073/pnas.89.12.5635. PMID 1376927.
- ↑ Coughlin P, Sun J, Cerruti L, Salem HH, Bird P., (1993). "Cloning and molecular characterization of a human intracellular serine proteinase inhibitor". Proc Natl Acad Sci U S A. 90 (20): 9417–21. doi:10.1073/pnas.90.20.9417. PMID 8415716.
- ↑ 28.0 28.1 Pak SC, Kumar V, Tsu C, Luke CJ, Askew YS, Askew DJ, Mills DR, Bromme D, Silverman GA. (2004). "SRP-2 is a cross-class inhibitor that participates in postembryonic development of the nematode Caenorhabditis elegans: initial characterization of the clade L serpins". J Biol Chem. 279 (15): 15448–59. doi:10.1074/jbc.M400261200. PMID 14739286.
- ↑ Bird PI. (1999). "Regulation of pro-apoptotic leucocyte granule serine proteinases by intracellular serpins". Immunol Cell Biol 77 (1): 47–57.. doi:10.1046/j.1440-1711.1999.00787.x. PMID 10101686.
- ↑ Bird CH, Sutton VR, Sun J, Hirst CE, Novak A, Kumar S, Trapani JA, Bird PI (1998). "Selective regulation of apoptosis: the cytotoxic lymphocyte serpin proteinase inhibitor 9 protects against granzyme B-mediated apoptosis without perturbing the Fas cell death pathway". Mol Cell Biol. 18 (11): 6387–98. PMID 774654..
- ↑ 31.0 31.1 Zou Z, Anisowicz A, Hendrix MJ, Thor A, Neveu M, Sheng S, Rafidi K, Seftor E, Sager R. (1994). "Maspin, a serpin with tumor-suppressing activity in human mammary epithelial cells". Science 263 (5146): 526–9. doi:10.1126/science.8290962. PMID 8290962.
- ↑ Luo JL, Tan W, Ricono JM, Korchynskyi O, Zhang M, Gonias SL, Cheresh DA, Karin M. (2007). "Nuclear cytokine-activated IKKalpha controls prostate cancer metastasis by repressing Maspin". Nature. 446 (7136): 690–4. doi:10.1038/nature05656. PMID 17377533.
- ↑ Grigoryev SA, Bednar J, Woodcock CL. (1999). "MENT, a heterochromatin protein that mediates higher order chromatin folding, is a new serpin family member". J Biol Chem. 274 (9): 5626–36. doi:10.1074/jbc.274.9.5626. PMID 10026180.
- ↑ Tasab M, Batten MR, Bulleid NJ (2000). "Hsp47: a molecular chaperone that interacts with and stabilizes correctly-folded procollagen". EMBO J. 19 (10): 2204–11. doi:10.1093/emboj/19.10.2204. PMID 10811611.
- ↑ Elliott PR, Lomas DA, Carrell RW, Abrahams JP. (1996). "Inhibitory conformation of the reactive loop of alpha 1-antitrypsin". Nat Struct Biol. 3 (8): 676–81. doi:10.1038/nsb0896-676. PMID 8756325.
- ↑ Horvath A, Irving J, Rossjohn J, Law R, Bottomley S, Quinsey N, Pike R, Coughlin P, Whisstock J (2005). "The murine orthologue of human antichymotrypsin: a structural paradigm for clade A3 serpins". J. Biol. Chem. 280 (52): 43168–78. doi:10.1074/jbc.M505598200. PMID 16141197.
- ↑ 37.0 37.1 Loebermann H, Tokuoka R, Deisenhofer J, Huber R. (1984). "Human alpha 1-proteinase inhibitor. Crystal structure analysis of two crystal modifications, molecular model and preliminary analysis of the implications for function". J Mol Biol. 177 (3): 531–57. doi:10.1016/0022-2836(84)90298-5. PMID 6332197.
- ↑ Stein PE, Leslie AG, Finch JT, Turnell WG, McLaughlin PJ, Carrell RW. (1990). "Crystal structure of ovalbumin as a model for the reactive centre of serpins". Nature. 347 (6288): 99–102. doi:10.1038/347099a0. PMID 2395463.
- ↑ 39.0 39.1 Whisstock J, Bottomley S (2006). "Molecular gymnastics: serpin structure, folding and misfolding". Curr Opin Struct Biol 16 (6): 761–8. doi:10.1016/j.sbi.2006.10.005. PMID 17079131.
- ↑ Egelund R, Rodenburg K, Andreasen P, Rasmussen M, Guldberg R, Petersen T (1998). "An ester bond linking a fragment of a serine proteinase to its serpin inhibitor". Biochemistry 37 (18): 6375–9. doi:10.1021/bi973043. PMID 9572853.
- ↑ Gettins P (2002). "Serpin structure, mechanism, and function". Chem Rev 102 (12): 4751–804. doi:10.1021/cr010170. PMID 12475206.
- ↑ Whisstock JC, Skinner R, Carrell RW, Lesk AM (2000). "Conformational changes in serpins: I. The native and cleaved conformations of alpha(1)-antitrypsin". J Mol Biol. 296 (2): 685–99. doi:10.1006/jmbi.1999.3520. PMID 10669617.
- ↑ Ye S, Cech A, Belmares R, Bergstrom R, Tong Y, Corey D, Kanost M, Goldsmith E (2001). "The structure of a Michaelis serpin-protease complex". Nat Struct Biol 8 (11): 979–83. doi:10.1038/nsb1101-979. PMID 11685246.
- ↑ Jin L, Abrahams JP, Skinner R, Petitou M, Pike RN, Carrell RW. (1997). "The anticoagulant activation of antithrombin by heparin". Proc Natl Acad Sci U S A. 94 (26): 14683–8. doi:10.1073/pnas.94.26.14683. PMID 9405673.
- ↑ Whisstock JC, Pike RN, Jin L, Skinner R, Pei XY, Carrell RW, Lesk AM. (2000). "Conformational changes in serpins: II. The mechanism of activation of antithrombin by heparin". J Mol Biol. 301 (5): 1287–305. doi:10.1006/jmbi.2000.3982. PMID 10966821.
- ↑ Li W, Johnson DJ, Esmon CT, Huntington JA (2004). "Structure of the antithrombin-thrombin-heparin ternary complex reveals the antithrombotic mechanism of heparin". Nat. Struct. Mol. Biol. 11 (9): 857–62. doi:10.1038/nsmb811. PMID 15311269.
- ↑ Johnson DJ, Li W, Adams TE, Huntington JA (2006). "Antithrombin-S195A factor Xa-heparin structure reveals the allosteric mechanism of antithrombin activation". EMBO J. 25 (9): 2029–37. doi:10.1038/sj.emboj.7601089. PMID 16619025.
- ↑ Petitou M, van Boeckel CA (2004). "A synthetic antithrombin III binding pentasaccharide is now a drug! What comes next?". Angew. Chem. Int. Ed. Engl. 43 (24): 3118–33. doi:10.1002/anie.200300640. PMID 15199558.
- ↑ Lindahl T, Sigurdardottir O, Wiman B (1989). "Stability of plasminogen activator inhibitor 1 (PAI-1)". Thromb Haemost 62 (2): 748–51. PMID 2479113.
- ↑ Zhang Q, Buckle AM, Law RH, Pearce MC, Cabrita LD, Lloyd GJ, Irving JA, Smith AI, Ruzyla K, Rossjohn J, Bottomley SP, Whisstock JC. (2007). "The N terminus of the serpin, tengpin, functions to trap the metastable native state". EMBO Rep. 8 (7): 658. doi:10.1038/sj.embor.7400986. PMID 17557112.
- ↑ Zhang Q, Law RH, Bottomley SP, Whisstock JC, Buckle AM (March 2008). "A structural basis for loop C-sheet polymerization in serpins". Journal of molecular biology 376 (5): 1348–59. doi:10.1016/j.jmb.2007.12.050. PMID 18234218.
- ↑ 52.0 52.1 Cao C, Lawrence DA, Li Y, Von Arnim CA, Herz J, Su EJ, Makarova A, Hyman BT, Strickland DK, Zhang L. (2006). "Endocytic receptor LRP together with tPA and PAI-1 coordinates Mac-1-dependent macrophage migration". EMBO J. 25 (9): 1860–70. doi:10.1038/sj.emboj.7601082. PMID 16601674.
- ↑ Jensen JK, Dolmer K, Gettins PG (May 2009). "Specificity of binding of the low density lipoprotein receptor-related protein (LRP) to different conformational states of the clade E serpins PAI-1 and PN1". J. Biol. Chem. 284 (27): 17989–97. doi:10.1074/jbc.M109.009530. PMID 19439404.
- ↑ Soukup SF, Culi J, Gubb D (June 2009). "Uptake of the Necrotic Serpin in Drosophila melanogaster via the Lipophorin Receptor-1". PLoS Genet. 5 (6): e1000532. doi:10.1371/journal.pgen.1000532. PMID 19557185.
- ↑ Pemberton PA, Stein PE, Pepys MB, Potter JM, Carrell RW (1988). "Hormone binding globulins undergo serpin conformational change in inflammation". Nature. 336 (6196): 257–8. doi:10.1038/336257a0. PMID 3143075.
- ↑ Chang WS, Whisstock J, Hopkins PC, Lesk AM, Carrell RW, Wardell MR. (1997). "Importance of the release of strand 1C to the polymerization mechanism of inhibitory serpins". Protein Sci. 6 (1): 89–98. doi:10.1002/pro.5560060110. PMID 9007980.
- ↑ Kroeger H, Miranda E, Macleod I, Perez J, Crowther DC, Marciniak SJ, Lomas DA (June 2009). "ERAD and autophagy cooperate to degrade polymerogenic mutant serpins". J. Biol. Chem. 284 (34): 22793–802. doi:10.1074/jbc.M109.027102. PMID 19549782.
- ↑ 58.0 58.1 Stein PE, Carrell RW. (1995). "What do dysfunctional serpins tell us about molecular mobility and disease?". Nat Struct Biol. 2 (2): 96–113.. doi:10.1038/nsb0295-96. PMID 7749926.
- ↑ 59.0 59.1 Huntington JA, Pannu NS, Hazes B, Read RJ, Lomas DA, Carrell RW (October 1999). "A 2.6 A structure of a serpin polymer and implications for conformational disease". Journal of molecular biology 293 (3): 449–55. doi:10.1006/jmbi.1999.3184. PMID 10543942.
- ↑ 60.0 60.1 Dunstone MA, Dai W, Whisstock JC, Rossjohn J, Pike RN, Feil SC, Le Bonniec BF, Parker MW & Bottomley SP (2000). "Cleaved antitrypsin polymers at atomic resolution". Protein Sci 9 (2): 417–420. doi:10.1110/ps.9.2.417. PMID 10716194.
- ↑ 61.0 61.1 61.2 Yu MH, Lee KN, Kim J (May 1995). "The Z type variation of human alpha 1-antitrypsin causes a protein folding defect". Nature structural biology 2 (5): 363–7. doi:10.1038/nsb0595-363. PMID 7664092.
- ↑ 62.0 62.1 62.2 Gooptu B, Hazes B, Chang WS, Dafforn TR, Carrell RW, Read RJ, Lomas DA. (2000). "Inactive conformation of the serpin alpha(1)-antichymotrypsin indicates two-stage insertion of the reactive loop: implications for inhibitory function and conformational disease". Proc Natl Acad Sci U S A. 97 (1): 67–72. doi:10.1073/pnas.97.1.67. PMID 10618372.
- ↑ Beauchamp NJ, Pike RN, Daly M, Butler L, Makris M, Dafforn TR, Zhou A, Fitton HL, Preston FE, Peake IR, Carrell RW (1998). "Antithrombins Wibble and Wobble (T85M/K): archetypal conformational diseases with in vivo latent-transition, thrombosis, and heparin activation". Blood 92 (8): 2696–706. PMID 9763552.
- ↑ Owen MC, Brennan SO, Lewis JH, Carrell RW. (1983). "Mutation of antitrypsin to antithrombin. alpha 1-antitrypsin Pittsburgh (358 Met leads to Arg), a fatal bleeding disorder". N Engl J Med. 309 (12): 694–8. doi:10.1056/NEJM198309223091203. PMID 6604220.
- ↑ Hopkins PC, Carrell RW, Stone SR. (1993). "Effects of mutations in the hinge region of serpins". Biochemistry. 32 (30): 7650–7. doi:10.1021/bi00081a008. PMID 8347575.
- ↑ 66.0 66.1 Gao F, Shi H, Daughty C, Cella N, Zhang M (2004). "Maspin plays an essential role in early embryonic development". Development 131 (7): 1479–89. doi:10.1242/dev.01048. PMID 14985257.
- ↑ Cabrita LD, Irving JA, Pearce MC, Whisstock JC, Bottomley SP. (2007). "Aeropin from the extremophile Pyrobaculum aerophilum bypasses the serpin misfolding trap". J Biol Chem. 282 (37): 26802. doi:10.1074/jbc.M705020200. PMID 17635906.
- ↑ Law RH, Zhang Q, McGowan S, Buckle AM, Silverman GA, Wong W, Rosado CJ, Langendorf CG, Pike RN, Bird PI, Whisstock JC (2006). "An overview of the serpin superfamily". Genome Biol. 7 (5): 216. doi:10.1186/gb-2006-7-5-216. PMID 16737556.
- ↑ Stoller JK, Aboussouan LS (2005). "Alpha1-antitrypsin deficiency". Lancet 365 (9478): 2225–36. doi:10.1016/S0140-6736(05)66781-5. PMID 15978931.
- ↑ Munch J, Standker L, Adermann K, Schulz A, Schindler M, Chinnadurai R, Pohlmann S, Chaipan C, Biet T, Peters T, Meyer B, Wilhelm D, Lu H, Jing W, Jiang S, Forssmann WG, Kirchhoff F. (2007). "Discovery and optimization of a natural HIV-1 entry inhibitor targeting the gp41 fusion peptide". Cell. 129 (2): 263–75. doi:10.1016/j.cell.2007.02.042. PMID 17448989.
- ↑ Seixas S, Suriano G, Carvalho F, Seruca R, Rocha J, Di Rienzo A. (2007). "Sequence Diversity at the Proximal 14q32.1 SERPIN Subcluster: Evidence for Natural Selection Favoring the Pseudogenization of SERPINA2". Mol Biol Evol. 24 (2): 587–98. doi:10.1093/molbev/msl187. PMID 17135331.
- ↑ Baumann U, Huber R, Bode W, Grosse D, Lesjak M, Laurell CB (April 1991). "Crystal structure of cleaved human alpha 1-antichymotrypsin at 2.7 A resolution and its comparison with other serpins". J. Mol. Biol. 218 (3): 595–606. doi:10.1016/0022-2836(91)90704-A. PMID 2016749.
- ↑ Kalsheker NA (1996). "Alpha 1-antichymotrypsin". Int. J. Biochem. Cell Biol. 28 (9): 961–4. PMID 8930118.
- ↑ Chao J, Stallone JN, Liang YM, Chen LM, Wang DZ, Chao L (July 1997). "Kallistatin is a potent new vasodilator". J. Clin. Invest. 100 (1): 11–7. doi:10.1172/JCI119502. PMID 9202051.
- ↑ Miao RQ, Agata J, Chao L, Chao J. (2002). "Kallistatin is a new inhibitor of angiogenesis and tumor growth". Blood 100 (9): 3245–52. doi:10.1182/blood-2002-01-0185. PMID 12384424.
- ↑ Li W, Adams TE, Kjellberg M, Stenflo J, Huntington JA. (2007). "Structure of native protein C inhibitor provides insight into its multiple functions". 282 282 (18): 13759–68. doi:10.1074/jbc.M701074200. PMID 17337440.
- ↑ Li W, Adams TE, Nangalia J, Esmon CT, Huntington JA. (2008). "Molecular basis of thrombin recognition by protein C inhibitor revealed by the 1.6-A structure of the heparin-bridged complex". Proc Natl Acad Sci U S A. 105 (12): 4661–6. doi:10.1073/pnas.0711055105. PMID 18362344.
- ↑ Geiger M (2007). "Protein C inhibitor, a serpin with functions in- and outside vascular biology". Thromb. Haemost. 97 (3): 343–7. PMID 17334499.
- ↑ Baumgärtner P, Geiger M, Zieseniss S, Malleier J, Huntington JA, Hochrainer K, Bielek E, Stoeckelhuber M, Lauber K, Scherfeld D, Schwille P, Wäldele K, Beyer K, Engelmann B (2007). "Phosphatidylethanolamine critically supports internalization of cell-penetrating protein C inhibitor". J. Cell Biol. 179 (4): 793–804. doi:10.1083/jcb.200707165. PMID 18025309.
- ↑ Uhrin P, Dewerchin M, Hilpert M, Chrenek P, Schofer C, Zechmeister-Machhart M, Kronke G, Vales A, Carmeliet P, Binder BR, Geiger M. (2000). "Disruption of the protein C inhibitor gene results in impaired spermatogenesis and male infertility". J Clin Invest. 106 (12): 1531–9. doi:10.1172/JCI10768. PMID 11120760.
- ↑ Han MH, Hwang SI, Roy DB, Lundgren DH, Price JV, Ousman SS, Fernald GH, Gerlitz B, Robinson WH, Baranzini SE, Grinnell BW, Raine CS, Sobel RA, Han DK, Steinman L (2008). "Proteomic analysis of active multiple sclerosis lesions reveals therapeutic targets". Nature 451 (7182): 1076. doi:10.1038/nature06559. PMID 18278032.
- ↑ Torpy DJ, Bachmann AW, Gartside M, Grice JE, Harris JM, Clifton P, Easteal S, Jackson RV, Whitworth JA. (2004). "Association between chronic fatigue syndrome and the corticosteroid-binding globulin gene ALA SER224 polymorphism". Endocr Res. 30 (3): 417–29. doi:10.1081/ERC-200035599. PMID 15554358.
- ↑ Bartalena L, Robbins J. (1992). "Variations in thyroid hormone transport proteins and their clinical implications". Thyroid. 2 (3): 237–45.. doi:10.1089/thy.1992.2.237. PMID 1422238.
- ↑ Kumar R, Singh VP, Baker KM (2007). "The intracellular renin-angiotensin system: a new paradigm". Trends Endocrinol. Metab. 18 (5): 208–14. doi:10.1016/j.tem.2007.05.001. PMID 17509892.
- ↑ Jeunemaitre X, Gimenez-Roqueplo AP, Celerier J, Corvol P. (1999). "Angiotensinogen variants and human hypertension". Curr Hypertens Rep. 1 (1): 31–41. doi:10.1007/s11906-999-0071-0. PMID 10981040.
- ↑ Tanimoto K, Sugiyama F, Goto Y, Ishida J, Takimoto E, Yagami K, Fukamizu A, Murakami K (1994). "Angiotensinogen-deficient mice with hypotension". J. Biol. Chem. 269 (50): 31334–7. PMID 7989296.
- ↑ Frazer JK, Jackson DG, Gaillard JP, Lutter M, Liu YJ, Banchereau J, Capra JD, Pascual V. (2000). "Identification of centerin: a novel human germinal center B cell-restricted serpin". Eur. J. Immunol. 30 (10): 3039–48. doi:10.1002/1521-4141(200010)30:10<3039::AID-IMMU3039>3.0.CO;2-H. PMID 11069088.
- ↑ Paterson MA, Horvath AJ, Pike RN, Coughlin PB. (2007). "Molecular characterization of centerin, a germinal centre cell serpin". Biochem J. 405 (3): 489–94. doi:10.1042/BJ20070174. PMID 17447896.
- ↑ Huang X, Dementiev A, Olson ST, Gettins PG (April 2010). "Basis for the specificity and activation of the serpin ZPI as an inhibitor of membrane-associated factor Xa". J Biol Chem 285 (26): 20399–409. doi:10.1074/jbc.M110.112748. PMID 20427285.
- ↑ Han X, Fiehler R, Broze GJ (2000). "Characterization of the protein Z-dependent protease inhibitor". Blood 96 (9): 3049–55. PMID 11049983.
- ↑ Corral J, Gonzalez-Conejero R, Soria JM, Gonzalez-Porras JR, Perez-Ceballos E, Lecumberri R, Roldan V, Souto JC, Minano A, Hernandez-Espinosa D, Alberca I, Fontcuberta J, Vicente V. (2006). "A nonsense polymorphism in the protein Z-dependent protease inhibitor increases the risk for venous thrombosis". Blood. 108 (1): 177–83. doi:10.1182/blood-2005-08-3249. PMID 16527896.
- ↑ Hida K, Wada J, Eguchi J, Zhang H, Baba M, Seida A, Hashimoto I, Okada T, Yasuhara A, Nakatsuka A, Shikata K, Hourai S, Futami J, Watanabe E, Matsuki Y, Hiramatsu R, Akagi S, Makino H, Kanwar YS (2005 J). "Visceral adipose tissue-derived serine protease inhibitor: a unique insulin-sensitizing adipocytokine in obesity". Proc. Natl. Acad. Sci. USA. 102 (30): 10610–5. doi:10.1073/pnas.0504703102. PMID 16030142.
- ↑ Baumann U, Bode W, Huber R, Travis J, Potempa J (August 1992). "Crystal structure of cleaved equine leucocyte elastase inhibitor determined at 1.95 A resolution". J. Mol. Biol. 226 (4): 1207–18. doi:10.1016/0022-2836(92)91062-T. PMID 1518052.
- ↑ Remold-O'Donnell E, Chin J, Alberts M. (1992). "Sequence and molecular characterization of human monocyte/neutrophil elastase inhibitor". Proc. Natl. Acad. Sci. USA. 89 (12): 5635–9. doi:10.1073/pnas.89.12.5635. PMID 1376927.
- ↑ Benarafa C, Priebe GP, Remold-O'donnell E. (2007). "The neutrophil serine protease inhibitor serpinb1 preserves lung defense functions in Pseudomonas aeruginosa infection". J. Exp. Med. 204 (8): 1901. doi:10.1084/jem.20070494. PMID 17664292.
- ↑ Harrop SJ, Jankova L, Coles M, Jardine D, Whittaker JS, Gould AR, Meister A, King GC, Mabbutt BC, Curmi PM (January 1999). "The crystal structure of plasminogen activator inhibitor 2 at 2.0 A resolution: implications for serpin function". Structure 7 (1): 43–54. doi:10.1016/S0969-2126(99)80008-2. PMID 10368272.
- ↑ Antalis TM, La Linn M, Donnan K, Mateo L, Gardner J, Dickinson JL, Buttigieg K, Suhrbier A (1998). "The serine proteinase inhibitor (serpin) plasminogen activation inhibitor type 2 protects against viral cytopathic effects by constitutive interferon alpha/beta priming". J. Exp. Med. 187 (11): 1799–811. doi:10.1084/jem.187.11.1799. PMID 9607921.
- ↑ Dougherty KM, Pearson JM, Yang AY, Westrick RJ, Baker MS, Ginsburg D. (1999). "The plasminogen activator inhibitor-2 gene is not required for normal murine development or survival". Proc Natl Acad Sci U S A. 96 (2): 686–91. doi:10.1073/pnas.96.2.686. PMID 9892694.
- ↑ Zheng B, Matoba Y, Kumagai T, Katagiri C, Hibino T, Sugiyama M (February 2009). "Crystal structure of SCCA1 and insight about the interaction with JNK1". Biochem. Biophys. Res. Commun. 380 (1): 143–7. doi:10.1016/j.bbrc.2009.01.057. PMID 19166818.
- ↑ Schick C, Kamachi Y, Bartuski AJ, Cataltepe S, Schechter NM, Pemberton PA, Silverman GA. (1997). "Squamous cell carcinoma antigen 2 is a novel serpin that inhibits the chymotrypsin-like proteinases cathepsin G and mast cell chymase". J Biol Chem. 272 (3): 1849–55. doi:10.1074/jbc.272.3.1849. PMID 8999871.
- ↑ Law RH, Irving JA, Buckle AM, Ruzyla K, Buzza M, Bashtannyk-Puhalovich TA, Beddoe TC, Nguyen K, Worrall DM, Bottomley SP, Bird PI, Rossjohn J, Whisstock JC (June 2005). "The high resolution crystal structure of the human tumor suppressor maspin reveals a novel conformational switch in the G-helix". J. Biol. Chem. 280 (23): 22356–64. doi:10.1074/jbc.M412043200. PMID 15760906.
- ↑ Teoh SS, Whisstock JC, Bird PI (April 2010). "Maspin (SERPINB5) is an obligate intracellular serpin". J. Biol. Chem. 285 (14): 10862–9. doi:10.1074/jbc.M109.073171. PMID 20123984.
- ↑ Scott FL, Hirst CE, Sun J, Bird CH, Bottomley SP, Bird PI (1999). "The intracellular serpin proteinase inhibitor 6 is expressed in monocytes and granulocytes and is a potent inhibitor of the azurophilic granule protease, cathepsin G". Blood 93 (6): 2089–97. PMID 10068683.
- ↑ Scarff KL, Ung KS, Nandurkar H, Crack PJ, Bird CH, Bird PI. (2004). "Targeted disruption of SPI3/Serpinb6 does not result in developmental or growth defects, leukocyte dysfunction, or susceptibility to stroke". Mol. Cell. Biol. 24 (9): 4075–82. doi:10.1128/MCB.24.9.4075-4082.2004. PMID 1508279.
- ↑ Sirmaci A, Erbek S, Price J, Huang M, Duman D, Cengiz FB, Bademci G, Tokgöz-Yilmaz S, Hişmi B, Ozdağ H, Oztürk B, Kulaksizoğlu S, Yildirim E, Kokotas H, Grigoriadou M, Petersen MB, Shahin H, Kanaan M, King MC, Chen ZY, Blanton SH, Liu XZ, Zuchner S, Akar N, Tekin M (May 2010). "A truncating mutation in SERPINB6 is associated with autosomal-recessive nonsyndromic sensorineural hearing loss". Am. J. Hum. Genet. 86 (5): 797–804. doi:10.1016/j.ajhg.2010.04.004. PMID 20451170.
- ↑ Miyata T, Inagi R, Nangaku M, Imasawa T, Sato M, Izuhara Y, Suzuki D, Yoshino A, Onogi H, Kimura M, Sugiyama S, Kurokawa K. (2002). "Overexpression of the serpin megsin induces progressive mesangial cell proliferation and expansion". J Clin Invest. 109 (5): 585–93. doi:10.1172/JCI14336. PMID 11877466.
- ↑ Dahlen JR, Jean F, Thomas G, Foster DC, Kisiel W (1998). "Inhibition of soluble recombinant furin by human proteinase inhibitor 8". J Biol Chem. 273 (4): 1851–4. doi:10.1074/jbc.273.4.1851. PMID 9442015.
- ↑ Sun J, Bird CH, Sutton V, McDonald L, Coughlin PB, De Jong TA, Trapani JA, Bird PI (1996). "A cytosolic granzyme B inhibitor related to the viral apoptotic regulator cytokine response modifier A is present in cytotoxic lymphocytes". J. Biol. Chem. 271 (44): 27802–9. doi:10.1074/jbc.271.44.27802. PMID 8910377.
- ↑ Zhang M, Park SM, Wang Y, Shah R, Liu N, Murmann AE, Wang CR, Peter ME, Ashton-Rickardt PG. (2006). "Serine protease inhibitor 6 protects cytotoxic T cells from self-inflicted injury by ensuring the integrity of cytotoxic granules". Immunity. 24 (4): 451–61. doi:10.1016/j.immuni.2006.02.002. PMID 16618603.
- ↑ Riewald M, Chuang T, Neubauer A, Riess H, Schleef RR. (1998). "Expression of bomapin, a novel human serpin, in normal/malignant hematopoiesis and in the monocytic cell lines THP-1 and AML-193". Blood. 9 (4): 1256–62. PMID 9454755.
- ↑ 111.0 111.1 Askew DJ, Cataltepe D, Kumar V, Edwards C, Pace SM, Howarth RN, Pak SC, Askew Y, Bromme D, Luke CJ, Whisstock JC, Silverman GA. (2007). "Serpinb11 is a new non-inhibitory intracellular serpin: Common single nucleotide polymorphisms in the scaffold impair conformational change". J. Biol. Chem. 282 (34): 24948. doi:10.1074/jbc.M703182200. PMID 17562709.
- ↑ Askew YS, Pak SC, Luke CJ, Askew DJ, Cataltepe S, Mills DR, Kato H, Lehoczky J, Dewar K, Birren B, Silverman GA. (2001). "SERPINB12 is a novel member of the human ov-serpin family that is widely expressed and inhibits trypsin-like serine proteinases". J. Biol. Chem. 276 (52): 49320–30. doi:10.1074/jbc.M108879200. PMID 11604408.
- ↑ Welss T, Sun J, Irving JA, Blum R, Smith AI, Whisstock JC, Pike RN, von Mikecz A, Ruzicka T, Bird PI, Abts HF. (2003). "Hurpin is a selective inhibitor of lysosomal cathepsin L and protects keratinocytes from ultraviolet-induced apoptosis". Biochemistry 42 (24): 7381–9. doi:10.1021/bi027307q. PMID 12809493.
- ↑ Huntington JA (2006). "Shape-shifting serpins--advantages of a mobile mechanism". Trends Biochem. Sci. 31 (8): 427–35. doi:10.1016/j.tibs.2006.06.005. PMID 16820297.
- ↑ Bruce D, Perry DJ, Borg JY, Carrell RW, Wardell MR. (1994). "Thromboembolic disease due to thermolabile conformational changes of antithrombin Rouen-VI (187 Asn-->Asp)". J. Clin. Invest. 94 (6): 2265–74. doi:10.1172/JCI117589. PMID 7989582.
- ↑ Baglin TP, Carrell RW, Church FC, Esmon CT, Huntington JA (2002). "Crystal structures of native and thrombin-complexed heparin cofactor II reveal a multistep allosteric mechanism". Proc. Natl. Acad. Sci. USA. 99 (17): 11079–84. doi:10.1073/pnas.162232399. PMID 12169660.
- ↑ Vicente CP, He L, Pavao MS, Tollefsen DM (2004). "Antithrombotic activity of dermatan sulfate in heparin cofactor II-deficient mice". Blood. 104 (13): 3965–70. doi:10.1182/blood-2004-02-0598. PMID 15315969.
- ↑ Aihara K, Azuma H, Akaike M, Ikeda Y, Sata M, Takamori N, Yagi S, Iwase T, Sumitomo Y, Kawano H, Yamada T, Fukuda T, Matsumoto T, Sekine K, Sato T, Nakamichi Y, Yamamoto Y, Yoshimura K, Watanabe T, Nakamura T, Oomizu A, Tsukada M, Hayashi H, Sudo T, Kato S, Matsumoto T. (2007). "Strain-dependent embryonic lethality and exaggerated vascular remodeling in heparin cofactor II-deficient mice". J. Clin. Invest. 117 (6): 1514–26. doi:10.1172/JCI27095. PMID 175492.
- ↑ Stout TJ, Graham H, Buckley DI, Matthews DJ (July 2000). "Structures of active and latent PAI-1: a possible stabilizing role for chloride ions". Biochemistry 39 (29): 8460–9. doi:10.1021/bi000290w. PMID 10913251.
- ↑ Zhou A, Huntington JA, Pannu NS, Carrell RW, Read RJ (July 2003). "How vitronectin binds PAI-1 to modulate fibrinolysis and cell migration". Nat. Struct. Biol. 10 (7): 541–4. doi:10.1038/nsb943. PMID 12808446.
- ↑ Cale JM, Lawrence DA (2007). "Structure-function relationships of plasminogen activator inhibitor-1 and its potential as a therapeutic agent". Curr. Drug Targets 8 (9): 971–81. doi:10.2174/138945007781662337. PMID 17896949.
- ↑ Gils A, Declerck PJ. (2004). "The structural basis for the pathophysiological relevance of PAI-I in cardiovascular diseases and the development of potential PAI-I inhibitors". Thromb. Haemost. 91 (3): 425–37. doi:10.1160/TH03-12-0764. PMID 14983217.
- ↑ Bajou K, Peng H, Laug WE, et al. (October 2008). "Plasminogen activator inhibitor-1 protects endothelial cells from FasL-mediated apoptosis". Cancer cell 14 (4): 324–34. doi:10.1016/j.ccr.2008.08.012. PMID 18835034.
- ↑ Lino MM, Atanasoski S, Kvajo M, Fayard B, Moreno E, Brenner HR, Suter U, Monard D (2007). "Mice lacking protease nexin-1 show delayed structural and functional recovery after sciatic nerve crush". J. Neurosci. 27 (14): 3677–85. doi:10.1523/JNEUROSCI.0277-07.2007. PMID 17409231.
- ↑ Murer V, Spetz JF, Hengst U, Altrogge LM, de Agostini A, Monard D. (2001). "Male fertility defects in mice lacking the serine protease inhibitor protease nexin-1". Proc. Natl. Acad. Sci. USA. 98 (6): 3029–33. doi:10.1073/pnas.051630698. PMID 11248026.
- ↑ Lüthi A, Van der Putten H, Botteri FM, Mansuy IM, Meins M, Frey U, Sansig G, Portet C, Schmutz M, Schröder M, Nitsch C, Laurent JP, Monard D (1997). "Endogenous serine protease inhibitor modulates epileptic activity and hippocampal long-term potentiation". J. Neurosci. 17 (12): 4688–99. PMID 9169529.
- ↑ Simonovic M, Gettins PG, Volz K (September 2001). "Crystal structure of human PEDF, a potent anti-angiogenic and neurite growth-promoting factor". Proc. Natl. Acad. Sci. U.S.A. 98 (20): 11131–5. doi:10.1073/pnas.211268598. PMID 11562499.
- ↑ 128.0 128.1 Doll JA, Stellmach VM, Bouck NP, Bergh AR, Lee C, Abramson LP, Cornwell ML, Pins MR, Borensztajn J, Crawford SE. (2003). "Pigment epithelium-derived factor regulates the vasculature and mass of the prostate and pancreas". Nat. Med. 9 (6): 774–80.. doi:10.1038/nm870. PMID 12740569.
- ↑ Becerra SP, Perez-Mediavilla LA, Weldon JE, et al. (September 2008). "pigment epithelium-derived factor (PEDF) binds to hyaluronan:Mapping of a hyaluronan binding site". The Journal of biological chemistry 283 (48): 33310. doi:10.1074/jbc.M801287200. PMID 18805795.
- ↑ PMID 19898467 (PubMed)
Citation will be completed automatically in a few minutes. Jump the queue or expand by hand - ↑ Law RH, Sofian T, Kan WT, Horvath AJ, Hitchen CR, Langendorf CG, Buckle AM, Whisstock JC, Coughlin PB (2008). "The X-ray crystal structure of the fibrinolysis inhibitor {alpha}2-antiplasmin". Blood 111 (4): 2049–2052. doi:10.1182/blood-2007-09-114215. PMID 18063751.
- ↑ Wiman B, Collen D (1979). "On the mechanism of the reaction between human alpha 2-antiplasmin and plasmin". J. Biol. Chem. 254 (18): 9291–7. PMID 158022.
- ↑ Miles LA, Plow EF, Donnelly KJ, Hougie C, Griffin JH. (1982). "A bleeding disorder due to deficiency of alpha 2-antiplasmin". Blood 59 (6): 1246–51. PMID 7082827.
- ↑ 134.0 134.1 Beinrohr L, Harmat V, Dobo J, Lorincz Z, Gal P, Zavodszky P. (2007). "C1-inhibitor serpin domain structure reveals the likely mechanism of heparin potentiation and conformational disease". J Biol Chem. 282 (29): 21100. doi:10.1074/jbc.M700841200. PMID 17488724.
- ↑ Aulak KS, Eldering E, Hack CE, Lubbers YP, Harrison RA, Mast A, Cicardi M, Davis AE 3rd. (1993). "A hinge region mutation in C1-inhibitor (Ala436-->Thr) results in nonsubstrate-like behavior and in polymerization of the molecule". J. Biol. Chem. 268 (24): 18088–94. PMID 8349686.
- ↑ Ennis S, Jomary C, Mullins R, et al. (October 2008). "Association between the SERPING1 gene and age-related macular degeneration: a two-stage case-control study". Lancet 372 (9652): 1828. doi:10.1016/S0140-6736(08)61348-3. PMID 18842294.
- ↑ Hirayoshi K, Kudo H, Takechi H, Nakai A, Iwamatsu A, Yamada KM, Nagata K (1991). "HSP47: a tissue-specific, transformation-sensitive, collagen-binding heat shock protein of chicken embryo fibroblasts". Mol. Cell. Biol. 11 (8): 4036–44. PMID 2072906.
- ↑ Nagai N, Hosokawa M, Itohara S, Adachi E, Matsushita T, Hosokawa N, Nagata K (2000). "Embryonic lethality of molecular chaperone hsp47 knockout mice is associated with defects in collagen biosynthesis". J Cell Biol. Year=2000 150 (6): 1499–506. doi:10.1083/jcb.150.6.1499. PMID 10995453.
- ↑ Christiansen HE, Schwarze U, Pyott SM, AlSwaid A, Al Balwi M, Alrasheed S, Pepin MG, Weis MA, Eyre DR, Byers PH (March 2010). "Homozygosity for a missense mutation in SERPINH1, which encodes the collagen chaperone protein HSP47, results in severe recessive osteogenesis imperfecta". Am. J. Hum. Genet. 86 (3): 389–98. doi:10.1016/j.ajhg.2010.01.034. PMID 20188343.
- ↑ Takehara S, Onda M, Zhang J, Nishiyama M, Yang X, Mikami B, Lomas DA (March 2009). "2.1A Crystal Structure of Native Neuroserpin Reveals Unique Structural Elements that Contribute to Conformational Instability". J. Mol. Biol. 388 (1): 11. doi:10.1016/j.jmb.2009.03.007. PMID 19285087.
- ↑ Ricagno S, Caccia S, Sorrentino G, Antonini G, Bolognesi M (March 2009). "Human Neuroserpin; Structure and Time-dependent Inhibition". J. Mol. Biol. 388 (1): 109. doi:10.1016/j.jmb.2009.02.056. PMID 19265707.
- ↑ Osterwalder T, Cinelli P, Baici A, Pennella A, Krueger SR, Schrimpf SP, Meins M, Sonderegger P (1998). "The axonally secreted serine proteinase inhibitor, neuroserpin, inhibits plasminogen activators and plasmin but not thrombin". J. Biol. Chem. 273 (4): 2312–21. doi:10.1074/jbc.273.4.2312. PMID 9442076.
- ↑ Davis RL, Shrimpton AE, Holohan PD, Bradshaw C, Feiglin D, Collins GH, Sonderegger P, Kinter J, Becker LM, Lacbawan F, Krasnewich D, Muenke M, Lawrence DA, Yerby MS, Shaw CM, Gooptu B, Elliott PR, Finch JT, Carrell RW, Lomas DA. (1999). "Familial dementia caused by polymerization of mutant neuroserpin". Nature 401 (6751): 376–9. doi:10.1038/43894. PMID 10517635.
- ↑ 144.0 144.1 Ozaki K, Nagata M, Suzuki M, Fujiwara T, Miyoshi Y, Ishikawa O, Ohigashi H, Imaoka S, Takahashi E, Nakamura Y. (1998). "Isolation and characterization of a novel human pancreas-specific gene, pancpin, that is down-regulated in pancreatic cancer cells". Genes Chromosomes and Cancer 22 (3): 179–85. doi:10.1002/(SICI)1098-2264(199807)22:3<179::AID-GCC3>3.0.CO;2-T. PMID 9624529.
- ↑ Loftus SK, Cannons JL, Incao A, Pak E, Chen A, Zerfas PM, Bryant MA, Biesecker LG, Schwartzberg PL, Pavan WJ (2005). "Acinar cell apoptosis in Serpini2-deficient mice models pancreatic insufficiency". PLoS. Genet. 1 (3): e38. doi:10.1371/journal.pgen.0010038. PMID 16184191.
- ↑ 146.0 146.1 Reichhart JM. (2005). "Tip of another iceberg: Drosophila serpins". Trends Cell Biol. 15 (12): 659–665. doi:10.1016/j.tcb.2005.10.001. PMID 16260136.
- ↑ Tang H, Kambris Z, Lemaitre B, Hashimoto C (October 2008). "A serpin that regulates immune melanization in the respiratory system of Drosophila". Developmental cell 15 (4): 617–26. doi:10.1016/j.devcel.2008.08.017. PMID 18854145.
- ↑ Scherfer C, Tang H, Kambris Z, Lhocine N, Hashimoto C, Lemaitre B (September 2008). "Drosophila Serpin-28D regulates hemolymph phenoloxidase activity and adult pigmentation". Developmental biology 323 (2): 189. doi:10.1016/j.ydbio.2008.08.030. PMID 18801354.
- ↑ Rushlow C (2004). "Dorsoventral patterning: a serpin pinned down at last". Curr. Biol. 14 (1): R16–8. doi:10.1016/j.cub.2003.12.015. PMID 14711428.
- ↑ Ligoxygakis P, Roth S, Reichhart JM (2003). "A serpin regulates dorsal-ventral axis formation in the Drosophila embryo". Curr. Biol. 13 (23): 2097–102. doi:10.1016/j.cub.2003.10.062. PMID 14654000.
- ↑ Hashimoto C, Kim DR, Weiss LA, Miller JW, Morisato D (2003). "Spatial regulation of developmental signaling by a serpin". Dev. Cell 5 (6): 945–50. doi:10.1016/S1534-5807(03)00338-1. PMID 14667416.
- ↑ Cliff J. Luke, Stephen C. Pak, Yuko S. Askew, Terra L. Naviglia, David J. Askew, Shila M. Nobar, Anne C. Vetica, Olivia S. Long, Simon C. Watkins, Donna B. Stolz, Robert J. Barstead, Gary L. Moulder, Dieter Brömme, and Gary A. Silverman (2007). "An Intracellular Serpin Regulates Necrosis by Inhibiting the Induction and Sequelae of Lysosomal Injury". Cell 130 (6): 1108–1119. doi:10.1016/j.cell.2007.07.013. PMID 17889653.
- ↑ Hejgaard J, Rasmussen SK, Brandt A, SvendsenI (1985). "Sequence homology between barley endosperm protein Z and protease inhibitors of the alpha-1-antitrypsin family". FEBS Lett. 180 (1): 89–94. doi:10.1016/0014-5793(85)80238-6.
- ↑ Dahl SW, Rasmussen SK, Peterson LC, Hejgaard J. (1996). "Inhibition of coagulation factors by recombinant barley serpin BSZx". FEBS Lett. 394 (2): 165–8. doi:10.1016/0014-5793(96)00940-4. PMID 8843156.
- ↑ Hejgaard J. (2001). "Inhibitory serpins from rye grain with glutamine as P1 and P2 residues in the reactive center". FEBS Lett. 488 (3): 149–53. doi:10.1016/S0014-5793(00)02425-X. PMID 11163762.
- ↑ Ostergaard H, Rasmussen SK, Roberts TH, Hejgaard J. (2000). "Inhibitory serpins from wheat grain with reactive centers resembling glutamine-rich repeats of prolamin storage proteins. Cloning and characterization of five major molecular forms". J Biol Chem. 275 (43): 33272–9. doi:10.1074/jbc.M004633200. PMID 10874043.
- ↑ 157.0 157.1 Yoo BC, Aoki K, Xiang Y, Campbell LR, Hull RJ, Xoconostle-Cázares B, Monzer J, Lee JY, Ullman DE, Lucas WJ. (2000). "Characterization of cucurbita maxima phloem serpin-1 (CmPS-1). A developmentally regulated elastase inhibitor". J Biol Chem. 275 (45): 35122–8. doi:10.1074/jbc.M006060200. PMID 10960478.
- ↑ la Cour Petersen M, Hejgaard J, Thompson GA, Schulz A. (2005). "Cucurbit phloem serpins are graft-transmissible and appear to be resistant to turnover in the sieve element-companion cell complex". J Exp Bot. 56 (422): 3111–20. doi:10.1093/jxb/eri308. PMID 16246856.
- ↑ Roberts TH, Hejgaard J. (2008). "Serpins in plants and green algae". Funct Integr Genomics. 8 (1): 1–27. doi:10.1007/s10142-007-0059-2. PMID 18060440.
- ↑ Vercammen D, Belenghi B, van de Cotte B, Beunens T, Gavigan JA, De Rycke R, Brackenier A, Inze D, Harris JL, Van Breusegem F. (2006). "Serpin1 of Arabidopsis thaliana is a suicide inhibitor for metacaspase 9". J Mol Biol. 364 (4): 625–36. doi:10.1016/j.jmb.2006.09.010. PMID 17028019.
- ↑ Ahn J-W, Atwell BJ, Roberts TH. (2009). "Serpin genes AtSRP2 and AtSRP3 are required for normal growth sensitivity to a DNA alkylating agent in Arabidopsis". BMC Plant Biol. 9: 52. doi:10.1186/1471-2229-9-52. PMID 19426562.
- ↑ 162.0 162.1 Kang S, Barak Y, Lamed R, Bayer EA, Morrison M. (2006). "The functional repertoire of prokaryote cellulosomes includes the serpin superfamily of serine proteinase inhibitors". Mol Microbiol. 60 (6): 1344–54. doi:10.1111/j.1365-2958.2006.05182.x. PMID 16796673.
- ↑ Irving JA, Cabrita LD, Rossjohn J, Pike RN, Bottomley SP, Whisstock JC (2003). "The 1.5 A crystal structure of a prokaryote serpin: controlling conformational change in a heated environment". Structure. 11 (4): 387–97.. doi:10.1016/S0969-2126(03)00057-1. PMID 12679017.
- ↑ Fulton KF, Buckle AM, Cabrita LD, Irving JA, Butcher RE, Smith I, Reeve S, Lesk AM, Bottomley SP, Rossjohn J, Whisstock JC. (2005). "The high resolution crystal structure of a native thermostable serpin reveals the complex mechanism underpinning the stressed to relaxed transition". J Biol Chem. 280 (9): 8435–42. doi:10.1074/jbc.M410206200. PMID 15590653.
- ↑ Turner PC, Moyer RW (2002). "Poxvirus immune modulators: functional insights from animal models". Virus Res. 88 (1-2): 35–53. doi:10.1016/S0168-1702(02)00119-3. PMID 12297326.
- ↑ 166.0 166.1 Richardson J, Viswanathan K, Lucas A (2006). "Serpins, the vasculature, and viral therapeutics". Front. Biosci. 11: 1042–56. doi:10.2741/1862. PMID 16146796.
- ↑ Jiang J, Arp J, Kubelik D, Zassoko R, Liu W, Wise Y, Macaulay C, Garcia B, McFadden G, Lucas AR, Wang H (2007). "Induction of indefinite cardiac allograft survival correlates with toll-like receptor 2 and 4 downregulation after serine protease inhibitor-1 (Serp-1) treatment" (Subscription required). Transplantation 84 (9): 1158–67. doi:10.1097/01.tp.0000286099.50532.b0. PMID 17998872.
- ↑ Dai E, Guan H, Liu L, Little S, McFadden G, Vaziri S, Cao H, Ivanova IA, Bocksch L, Lucas A (2003). "Serp-1, a viral anti-inflammatory serpin, regulates cellular serine proteinase and serpin responses to vascular injury". J. Biol. Chem. 278 (20): 18563–72. doi:10.1074/jbc.M209683200. PMID 12637546.
- ↑ Turner PC, Sancho MC, Thoennes SR, Caputo A, Bleackley RC, Moyer RW (1999). "Myxoma virus Serp2 is a weak inhibitor of granzyme B and interleukin-1beta-converting enzyme in vitro and unlike CrmA cannot block apoptosis in cowpox virus-infected cells". J. Virol. 73 (8): 6394–404. PMID 10400732.
- ↑ Munuswamy-Ramanujam G, Khan KA, Lucas AR (2006). "Viral anti-inflammatory reagents: the potential for treatment of arthritic and vasculitic disorders". Endocr Metab Immune Disord Drug Targets 6 (4): 331–43. PMID 17214579.
- ↑ Renatus M, Zhou Q, Stennicke HR, Snipas SJ, Turk D, Bankston LA, Liddington RC, Salvesen GS (2000). "Crystal structure of the apoptotic suppressor CrmA in its cleaved form". Structure 8 (7): 789–97. doi:10.1016/S0969-2126(00)00165-9. PMID 10903953.
External links
- PDB Molecule of the Month pdb53_1
- MeSH Serpins
- James Whisstock laboratory at Monash University
- Jim Huntington serpin laboratory at University of Cambridge
- Frank Church serpin group at University of North Carolina at Chapel Hill
- Paul Declerck serpin group at Katholieke Universiteit Leuven
- Merops general protease / inhibitor classification page at University of Cambridge
- The alpha one foundation provides information and support for sufferers of antitrypsin deficiency
|
|||||||||||