Papillomaviridae
| Papillomavirus | |
|---|---|
 |
|
| EM of papillomavirus | |
| Virus classification | |
| Group: | Group I (dsDNA) |
| Family: | Papillomaviridae |
| Genera | |
|
Alphapapillomavirus |
|
Papillomaviridae is an ancient and diverse taxonomic family of non-enveloped DNA viruses, collectively known as papillomaviruses. Several hundred species of papillomaviruses, traditionally referred to as "types",[1] have been identified infecting all carefully inspected birds and mammals,[1] but also a small number of reptiles, in particular turtles and tortoises.[2][3] Infection by most papillomavirus types, depending on the type, is either asymptomatic or causes small benign tumors, known as papillomas or warts. Papillomas caused by some types, however, such as human papillomaviruses 16 and 18, carry a risk of becoming cancerous.[4]
Papillomaviruses are highly host- and tissue-tropic, and are rarely transmitted between species.[5] Papillomaviruses replicate exclusively in the basal layer of the body surface tissues. All known papillomavirus types infect a particular body surface,[1] typically the skin or mucosal epithelium of the genitals, anus, mouth, or airways.[6] For example, human papillomavirus (HPV) type 1 tends to infect the soles of the feet, and HPV type 2 the palms of the hands, where they may cause warts.
Papillomaviruses were first identified in the early 20th century, when it was shown that skin warts, or papillomas, could be transmitted between individuals by a filterable infectious agent. In 1935 Francis Peyton Rous, who had previously demonstrated the existence of a cancer-causing sarcoma virus in chickens, went on to show that a papillomavirus could cause skin cancer in infected rabbits. This was the first demonstration that a virus could cause cancer in mammals.
Contents |
Taxonomy of papillomaviruses

Papillomaviruses (PVs) have similar genomic organizations, and any pair of PVs contains at least five homologous genes, although the nucleotide sequence may diverge by more than 50%. Phylogenetic algorithms that permit the comparison of homologies led to phylogenetic trees that have a similar topology, independent of the gene analyzed. Phylogenetic studies strongly suggest that PVs normally evolve together with their mammalian and bird host species, do not change host species, do not recombine, and have maintained their basic genomic organization for a period exceeding 100 million years. These sequence comparisons have laid the foundation for a PV taxonomy, which is now officially recognized by the International Committee on Taxonomy of Viruses. All PVs form the family Papillomaviridae, which is distinct from the Polyomaviridae thus eliminating the term Papovaviridae. Major branches of the phylogenetic tree of PVs are considered genera, which are identified by Greek letters. Minor branches are considered species and unite PV types that are genomically distinct without exhibiting known biological differences. This new taxonomic system does not affect the traditional identification and characterization of PV "types" and their independent isolates with minor genomic differences, referred to as "subtypes" and "variants", all of which are taxa below the level of "species".[7]
Animal papillomaviruses
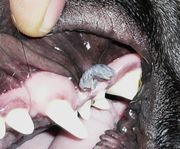
Individual papillomavirus types tend to be highly adapted to replication in a single animal species. In one study, researchers swabbed the forehead skin of a variety of zoo animals and used PCR to amplify any papillomavirus DNA that might be present.[8] Although a wide variety of papillomavirus sequences were identified in the study, the authors found little evidence for inter-species transmission. Interestingly, one zookeeper was found to be transiently positive for a chimpanzee-specific papillomavirus sequence. However, the authors note that the chimpanzee-specific papillomavirus sequence could have been the result of surface contamination of the zookeeper's skin, as opposed to productive infection.
Cottontail rabbit papillomavirus (CRPV) can cause protuberant warts (see images) in its native host, the North American rabbit genus Sylvilagus. These horn-like warts may be the original basis for the urban legends of the American antlered rabbit the Jackalope and European Wolpertinger. European domestic rabbits (genus Oryctolagus) can be transiently infected with CRPV in a laboratory setting. However, since European domestic rabbits do not produce infectious progeny virus, they are considered an incidental or "dead-end" host for CRPV.[9]
Inter-species transmission has also been documented for bovine papillomavirus (BPV) type 1.[10] In its natural host (cattle), BPV-1 induces large fibrous skin warts. BPV-1 infection of horses, which are an incidental host for the virus, can lead to the development of benign tumors known as sarcoids. The agricultural significance of BPV-1 spurred a successful effort to develop a vaccine against the virus.
A few reports have identified papillomaviruses in smaller rodents, such as Syrian hamsters, the African multimammate rat and the European harvest mouse.[11] However, there are no papillomaviruses known to be capable of infecting laboratory mice. The lack of a tractable mouse model for papillomavirus infection has been a major limitation for laboratory investigation of papillomaviruses.
Evolution
The evolution of papillomaviruses is slow compared to many other virus types. This is probably because the papillomavirus genome is composed of genetically stable double-stranded DNA that is replicated with high fidelity by the host cell's DNA replication machinery.
It is believed that papillomaviruses generally co-evolve with a particular species of host animal over many years. In a particularly speedy example, HPV-16 has evolved slightly as human populations have expanded across the globe and now varies in different geographic regions in a way that probably reflects the history of human migration.[12][13]
Other HPV types, such as HPV-13, vary relatively little in different human populations. In fact, the sequence of HPV-13 closely resembles a papillomavirus of bonobos (also known as pygmy chimpanzees).[14] It is not clear whether this similarity is due to recent transmission between species or because HPV-13 has simply changed very little in the six or so million years since humans and bonobos diverged.[13]
Structure

Papillomaviruses are non-enveloped, meaning that the outer shell or capsid of the virus is not covered by a lipid membrane. A single viral protein, known as L1, is necessary and sufficient for formation of a 60 nanometer capsid composed of 72 star-shaped capsomers (see figure). Like most non-enveloped viruses, the capsid is geometrically regular and presents icosahedral symmetry. Self-assembled virus-like particles composed of L1 are the basis of a successful group of prophylactic HPV vaccines designed to elicit virus-neutralizing antibodies that protect against initial HPV infection.
The papillomavirus genome is a double-stranded circular DNA molecule ~8,000 base pairs in length. It is packaged within the L1 shell along with cellular histone proteins, which serve to wrap and condense DNA.
The papillomavirus capsid also contains a viral protein known as L2, which is less abundant. Although not clear how L2 is arranged within the virion, it is known to perform several important functions, including facilitating the packaging of the viral genome into nascent virions as well as the infectious entry of the virus into new host cells. L2 is of interest as a possible target for more broadly protective HPV vaccines.
Tissue specificity
Papillomaviruses replicate exclusively in keratinocytes. Keratinocytes form the outermost layers of the skin, as well as some mucosal surfaces, such as the inside of the cheek or the walls of the vagina. These surface tissues, which are known as stratified squamous epithelia, are composed of stacked layers of flattened cells. The cell layers are formed through a process known as cellular differentiation, in which keratinocytes gradually become specialized, eventually forming a hard, crosslinked surface that prevents moisture loss and acts as a barrier against pathogens. Less-differentiated keratinocyte stem cells, replenished on the surface layer, are thought to be the initial target of productive papillomavirus infections. Subsequent steps in the viral life cycle are strictly dependent on the process of keratinocyte differentiation. As a result, papillomaviruses can only replicate in body surface tissues.
Life cycle
Infectious entry
Papillomaviruses gain access to keratinocyte stem cells through small wounds, known as microtraumas, in the skin or mucosal surface. Interactions between L1 and sulfated sugars on the cell surface promote initial attachment of the virus.[15][16] The virus is then able to get inside from the cell surface via interaction with a specific receptor, likely via the alpha-6 beta-4 integrin,[17][18] and transported to membrane-enclosed vesicles called endosomes.[19][20] The capsid protein L2 disrupts the membrane of the endosome, allowing the viral genome to escape and traffic, along with L2, to the cell nucleus.[21][22]
Viral Persistence
After successful infection of a keratinocyte, the virus expresses E1 and E2 proteins, which are for replicating and maintaining the viral DNA as a circular episome. The viral oncogenes E6 and E7 promote cell growth by inactivating the tumor suppressor proteins p53 and pRb. Keratinocyte stem cells in the epithelial basement layer can maintain papillomavirus genomes for decades.[6]
Production of progeny virus
The expression of the viral late genes, L1 and L2, is exclusively restricted to differentiating keratinocytes in the outermost layers of the skin or mucosal surface. The increased expression of L1 and L2 is typically correlated with a dramatic increase in the number of copies of the viral genome. Since the outer layers of stratified squamous epithelia are subject to relatively limited surveillance by cells of the immune system, it is thought that this restriction of viral late gene expression represents a form of immune evasion.
New infectious progeny virus are assembled in the cell nucleus. Papillomaviruses have evolved a mechanism for releasing virions into the environment. Other kinds of non-enveloped animal viruses utilize an active lytic process to kill the host cell, allowing release of progeny virus particles. Often this lytic process is associated with inflammation, which might trigger immune attack against the virus. Papillomaviruses exploit desquamation as a stealthy, non-inflammatory release mechanism.
Cancer
Although some papillomavirus types can cause cancer in the epithelial tissues they inhabit, cancer is not a typical outcome of infection. The development of papillomavirus-induced cancers typically occurs over the course of many years.
Laboratory study
The fact that the papillomavirus life cycle strictly requires keratinocyte differentiation has posed a substantial barrier to the study of papillomaviruses in the laboratory, since it has precluded the use of conventional cell lines to grow the viruses. Because infectious BPV-1 virions can be extracted from the large warts the virus induces on cattle, it has been a workhorse model papillomavirus type for many years. CRPV, rabbit oral papillomavirus (ROPV) and canine oral papillomavirus (COPV) have also been used extensively for laboratory studies.
Some sexually transmitted HPV types have been propagated using a mouse “xenograft” system, in which HPV-infected human cells are implanted into immunodeficient mice. More recently, some groups have succeeded in isolating infectious HPV-16 from human cervical lesions. However, isolation of infectious virions using this technique is arduous and the yield of infectious virus is very low.
The differentiation of keratinocytes can be mimicked in vitro by exposing cultured keratinocytes to an air/liquid interface. The adaptation of such “raft culture” systems to the study of papillomaviruses was a significant breakthrough for in vitro study of the viral life cycle.[23] However, raft culture systems are relatively cumbersome and the yield of infectious HPVs can be low.[24]
The development of a yeast-based system that allows stable episomal HPV replication provides a convenient, rapid and inexpensive means to study several aspects of the HPV lifecycle (Angeletti 2002). For example, E2-dependent transcription, genome amplification and efficient encapsidation of full-length HPV DNAs can be easily recreated in yeast (Angeletti 2005).
Recently, transient high-yield methods for producing HPV pseudoviruses carrying reporter genes has been developed. Although pseudoviruses are not suitable for studying certain aspects of the viral life cycle, initial studies suggest that their structure and initial infectious entry into cells is probably similar in many ways to authentic papillomaviruses.
Genetic organization

The papillomavirus genome is divided into an early region (E), encoding various genes that are expressed immediately after initial infection of a host cell, and a late region (L) encoding the capsid genes L1 and L2. All the genes are encoded on one DNA strand (see figure). This represents a dramatic difference between papillomaviruses and polyomaviruses, since the latter virus type expresses its early and late genes by bi-directional transcription of both DNA strands. This difference was a major factor in establishment of the consensus that papillomaviruses and polyomaviruses probably never shared a common ancestor, despite the striking similarities in the structures of their virions.
Technical discussion of papillomavirus gene functions
Genes within the papillomavirus genome are usually identified after similarity with other previously identified genes. However, some spurious open reading frames might have been mistaken as genes simply after their position in the genome, and might not be true genes. This applies specially to certain E3, E4, E5 and E8 open reading frames.
E1
Encodes a protein that binds to the viral origin of replication in the long control region of the viral genome. E1 uses ATP to exert a helicase activity that forces apart the DNA strands, thus preparing the viral genome for replication by cellular DNA replication factors.
E2
The E2 protein serves as a master transcriptional regulator for viral promoters located primarily in the long control region. The protein has a transactivation domain linked by a relatively unstructured hinge region to a well-characterized DNA binding domain. E2 facilitates the binding of E1 to the viral origin of replication. E2 also utilizes a cellular protein known as Bromodomain-4 (Brd4) to tether the viral genome to cellular chromosomes.[25] This tethering to the cell's nuclear matrix ensures faithful distribution of viral genomes to each daughter cell after cell division. It is thought that E2 serves as a negative regulator of expression for the oncogenes E6 and E7 in latently HPV-infected basal layer keratinocytes. Genetic changes, such as integration of the viral DNA into a host cell chromosome, that inactivate E2 expression tend to increase the expression of the E6 and E7 oncogenes, resulting in cellular transformation and possibly further genetic destabilization.
E3
This small putative gene exists only in a few papillomavirus types. The gene is not known to be expressed as a protein and does not appear to serve any function.
E4
Although E4 proteins are expressed at low levels during the early phase of viral infection, expression of E4 increases dramatically during the late phase of infection. In other words, its “E” appellation may be something of a misnomer. In the case of HPV-1, E4 can account for up to 30% of the total protein at the surface of a wart.[26] The E4 protein of many papillomavirus types is thought to facilitate virion release into the environment by disrupting intermediate filaments of the keratinocyte cytoskeleton. Viral mutants incapable of expressing E4 do not support high-level replication of the viral DNA, but it is not yet clear how E4 facilitates DNA replication. E4 has also been shown to participate in arresting cells in the G2 phase of the cell cycle.
E5
The E5 are small, very hydrophobic proteins that destabilise the function of many membrane proteins in the infected cell.[27] The E5 protein of some animal papillomavirus types (mainly bovine papillomavirus type 1) functions as an oncogene primarily by activating the cell growth-promoting signaling of platelet-derived growth factor receptors. The E5 proteins of human papillomaviruses associated to cancer, however, seem to activate the signal cascade initiated by epidermal growth factor upon ligand binding. HPV16 E5 and HPV2 E5 have also been shown to down-regulate the surface expression of major histocompatibility complex class I proteins, which may prevent the infected cell from being eliminated by killer T cells.
E6

E6 is a 151 amino-acid peptide that incorporates a type 1 motif with a consensus sequence –(T/S)-(X)-(V/I)-COOH.[28][29] It also has two zinc finger motifs.[28]
E6 is of particular interest because it appears to have multiple roles in the cell and to interact with many other proteins. Its major role, however, is to mediate the degradation of p53, a major tumor suppressor protein, reducing the cell's ability to respond to DNA damage. [30][31]
E6 has also been shown to target other cellular proteins, thereby altering several metabolic pathways. One such target is NFX1-91, which normally represses production of telomerase, a protein that allows cells to divide an unlimited number of times. When NFX1-91 is degraded by E6, telomerase levels increase, inactivating a major mechanism keeping cell growth in check.[32] Additionally, E6 can act as a transcriptional cofactor—specifically, a transcription activator—when interacting with the cellular transcription factor, E2F1/DP1.[28]
E6 can also bind to PDZ-domains, short sequences which are often found in signaling proteins. E6's structural motif allows for interaction with PDZ domains on DLG (discs large) and hDLG (Drosophila large) tumor suppressor genes.[29][33] Binding at these locations causes transformation of the DLG protein and disruption of its suppressor function. E6 proteins also interact with the MAGUK (membrane-associated guanylate kinase family) proteins. These proteins, including MAGI-1, MAGI-2, and MAGI-3 are usually structural proteins, and can help with signaling.[29][33] More significantly, they are believed to be involved with DLG's suppression activity. When E6 complexes with the PDZ domains on the MAGI proteins, it distorts their shape and thereby impedes their function. Overall, the E6 protein serves to impede normal protein activity in such a way as to allow a cell to grow and multiply at the increased rate characteristic of cancer.
Since the expression of E6 is strictly required for maintenance of a malignant phenotype in HPV-induced cancers, it is an appealing target of therapeutic HPV vaccines designed to eradicate established cervical cancer tumors.
E7
In most papillomavirus types, the primary function of the E7 protein is to inactivate members of the pRb family of tumor suppressor proteins. Together with E6, E7 serves to prevent cell death (apoptosis) and promote cell cycle progression, thus priming the cell for replication of the viral DNA. E7 also participates in immortalization of infected cells by activating cellular telomerase. Like E6, E7 is the subject of intense research interest and is believed to exert a wide variety of other effects on infected cells. As with E6, the ongoing expression of E7 is required for survival of cancer cell lines, such as HeLa, that are derived from HPV-induced tumors.[34]
E8
Only a few papillomavirus types encode a short protein from the E8 gene. In the case of BPV-4 (papillomavirus genus Xi), the E8 open reading frame may substitute for the E6 open reading frame, which is absent in this papillomavirus genus.[35] These E8 genes are chemically and functionally similar to the E5 genes from some human papillomaviruses, and are also called E5/E8.
L1
L1 spontaneously self-assembles into pentameric capsomers. Purified capsomers can go on to form capsids, which are stabilized by disulfide bonds between neighboring L1 molecules. L1 capsids assembled in vitro are the basis of prophylactic vaccines against several HPV types. Compared to other papillomavirus genes, the amino acid sequences of most portions of L1 are well-conserved between types. However, the surface loops of L1 can differ substantially, even for different members of a particular papillomavirus species. This probably reflects a mechanism for evasion of neutralizing antibody responses elicited by previous papillomavirus infections.[36]
L2
L2 exists in an oxidized state within the papillomavirus virion, with the two conserved cysteine residues forming an intramolecular disulfide bond.[37] In addition to cooperating with L1 to package the viral DNA into the virion, L2 has been shown to interact with a number of cellular proteins during the infectious entry process. After the initial binding of the virion to the cell, L2 must be cleaved by the cellular protease furin.[38] The virion is internalized, probably through a clathrin-mediated process, into an endosome, where acidic conditions are thought to lead to exposure of membrane-destabilizing portions of L2.[21] The cellular proteins beta-actin[39] and syntaxin-18[40] may also participate in L2-mediated entry events. After endosome escape, L2 and the viral genome are imported into the cell nucleus where they traffic to a sub-nuclear domain known as an ND-10 body that is rich in transcription factors.[22] Small portions of L2 are well-conserved between different papillomavirus types, and experimental vaccines targeting these conserved domains may offer protection against a broad range of HPV types.[41]
External links
- Viralzone: Papillomaviridae
- Los Alamos National Laboratory maintains a comprehensive (albeit somewhat dated) papillomavirus sequence database. This useful database provides detailed descriptions and references for various papillomavirus types.
- A short video which shows the effects of papillomavirus on the skin of an Indonesian man with epidermodysplasia verruciformis, the genetic inability to defend against some types of cutaneous HPV.
- [1] de Villiers, E.M., Bernard, H.U., Broker, T., Delius, H. and zur Hausen, H. Index of Viruses - Papillomaviridae (2006). In: ICTVdB - The Universal Virus Database, version 4. Büchen-Osmond, C (Ed), Columbia University, New York, USA.
- 00.099. Papillomaviridae description In: ICTVdB - The Universal Virus Database, version 4. Büchen-Osmond, C. (Ed), Columbia University, New York, USA
References
- ↑ 1.0 1.1 1.2 de Villiers EM, Fauquet C, Broker TR, Bernard HU, zur Hausen H. (2004). "Classification of papillomaviruses". Virology 324 (1): 17–27. doi:10.1016/j.virol.2004.03.033. PMID 15183049.
- ↑ Herbst LH, Lenz J, Van Doorslaer K, Chen Z, Stacy BA, Wellehan JF Jr, Manire CA, Burk RD (2009). "Genomic characterization of two novel reptilian papillomaviruses, Chelonia mydas papillomavirus 1 and Caretta caretta papillomavirus 1". Virology 383 (1): 131–5. doi:10.1016/j.virol.2008.09.022. PMID 18973915.
- ↑ Drury SE, Gough RE, McArthur S, Jessop M (1998). "Detection of herpesvirus-like and papillomavirus-like particles associated with diseases of tortoises". Vet Rec. 143 (23): 639. PMID 9881444.
- ↑ Muñoza N, Castellsaguéb X, Berrington de Gonzálezc A, Gissmann L (2006). "Chapter 1: HPV in the etiology of human cancer". Vaccine 24 (3): S1–S10. doi:10.1016/j.vaccine.2006.05.115. PMID 16949995.
- ↑ Mistry N, Wibom C, Evander M (2008). "Cutaneous and mucosal human papillomaviruses differ in net surface charge, potential impact on tropism". Virol J. 5: 118. doi:10.1186/1743-422X-5-118. PMID 18854037.
- ↑ 6.0 6.1 Doorbar J (2005). "The papillomavirus life cycle". J. Clin. Virol. 32 Suppl 1: S7–15. doi:10.1016/j.jcv.2004.12.006. PMID 15753007.
- ↑ Campo MS (editor). (2006). Papillomavirus Research: From Natural History To Vaccines and Beyond. Caister Academic Press. ISBN 978-1-904455-04-2 . http://www.horizonpress.com/pv.
- ↑ Antonsson A, Hansson BG (2002). "Healthy skin of many animal species harbors papillomaviruses which are closely related to their human counterparts". J. Virol. 76 (24): 12537–42. doi:10.1128/JVI.76.24.12537-12542.2002. PMID 12438579.
- ↑ Christensen ND (2005). "Cottontail rabbit papillomavirus (CRPV) model system to test antiviral and immunotherapeutic strategies". Antivir. Chem. Chemother. 16 (6): 355–62. PMID 16331841.
- ↑ Coggins LW, Ma JQ, Slater AA, Campo MS (1985). "Sequence homologies between bovine papillomavirus genomes mapped by a novel low-stringency heteroduplex method". Virology 143 (2): 603–11. doi:10.1016/0042-6822(85)90398-8. PMID 2998027.
- ↑ Van Ranst M, Tachezy R, Pruss J, Burk RD (1992). "Primary structure of the E6 protein of Micromys minutus papillomavirus and Mastomys natalensis papillomavirus". Nucleic Acids Res. 20 (11): 2889. doi:10.1093/nar/20.11.2889. PMID 1319576.
- ↑ Ho L, Chan SY, Burk RD, et al. (1993). "The genetic drift of human papillomavirus type 16 is a means of reconstructing prehistoric viral spread and the movement of ancient human populations". J. Virol. 67 (11): 6413–23. PMID 8411343.
- ↑ 13.0 13.1 Calleja-Macias IE, Villa LL, Prado JC, et al. (2005). "Worldwide genomic diversity of the high-risk human papillomavirus types 31, 35, 52, and 58, four close relatives of human papillomavirus type 16". J. Virol. 79 (21): 13630–40. doi:10.1128/JVI.79.21.13630-13640.2005. PMID 16227283.
- ↑ Van Ranst M, Fuse A, Fiten P, et al. (1992). "Human papillomavirus type 13 and pygmy chimpanzee papillomavirus type 1: comparison of the genome organizations". Virology 190 (2): 587–96. doi:10.1016/0042-6822(92)90896-W. PMID 1325697.
- ↑ Joyce JG, Tung JS, Przysiecki CT, et al. (1999). "The L1 major capsid protein of human papillomavirus type 11 recombinant virus-like particles interacts with heparin and cell-surface glycosaminoglycans on human keratinocytes". J. Biol. Chem. 274 (9): 5810–22. doi:10.1074/jbc.274.9.5810. PMID 10026203.
- ↑ Giroglou T, Florin L, Schäfer F, Streeck RE, Sapp M (2001). "Human papillomavirus infection requires cell surface heparan sulfate". J. Virol. 75 (3): 1565–70. doi:10.1128/JVI.75.3.1565-1570.2001. PMID 11152531.
- ↑ Evander M, Frazer IH, Payne E, Qi YM, Hengst K, McMillan NA (1997). "Identification of the alpha6 integrin as a candidate receptor for papillomaviruses". J. Virol. 71 (3): 2449–56. PMID 9032382.
- ↑ McMillan NA, Payne E, Frazer IH, Evander M (1999). "Expression of the alpha6 integrin confers papillomavirus binding upon receptor-negative B-cells". Virology 261 (2): 271–9. doi:10.1006/viro.1999.9825. PMID 10497112.
- ↑ Selinka HC, Giroglou T, Sapp M (2002). "Analysis of the infectious entry pathway of human papillomavirus type 33 pseudovirions". Virology 299 (2): 279–287. doi:10.1006/viro.2001.1493. PMID 12202231.
- ↑ Day PM, Lowy DR, Schiller JT (2003). "Papillomaviruses infect cells via a clathrin-dependent pathway". Virology 307 (1): 1–11. doi:10.1016/S0042-6822(02)00143-5. PMID 12667809.
- ↑ 21.0 21.1 Kämper N, Day PM, Nowak T, et al. (2006). "A membrane-destabilizing peptide in capsid protein L2 is required for egress of papillomavirus genomes from endosomes". J. Virol. 80 (2): 759–68. doi:10.1128/JVI.80.2.759-768.2006. PMID 16378978.
- ↑ 22.0 22.1 Day PM, Baker CC, Lowy DR, Schiller JT (2004). "Establishment of papillomavirus infection is enhanced by promyelocytic leukemia protein (PML) expression". Proc. Natl. Acad. Sci. U.S.A. 101 (39): 14252–7. doi:10.1073/pnas.0404229101. PMID 15383670.
- ↑ Meyers C, Frattini MG, Hudson JB, Laimins LA (1992). "Biosynthesis of human papillomavirus from a continuous cell line upon epithelial differentiation". Science 257 (5072): 971–3. doi:10.1126/science.1323879. PMID 1323879.
- ↑ McLaughlin-Drubin ME, Christensen ND, Meyers C (2004). "Propagation, infection, and neutralization of authentic HPV16 virus". Virology 322 (2): 213–9. doi:10.1016/j.virol.2004.02.011. PMID 15110519.
- ↑ McBride AA, McPhillips MG, Oliveira JG (2004). "Brd4: tethering, segregation and beyond". Trends Microbiol. 12 (12): 527–9. doi:10.1016/j.tim.2004.10.002. PMID 15539109.
- ↑ Doorbar J, Campbell D, Grand RJ, Gallimore PH (1986). "Identification of the human papilloma virus-1a E4 gene products". EMBO J. 5 (2): 355–62. PMID 3011404.
- ↑ Bravo IG, Alonso A (2004). "Mucosal human papillomaviruses encode four different E5 proteins whose chemistry and phylogeny correlate with malignant or benign growth". J. Virol. 78 (24): 13613–26. doi:10.1128/JVI.78.24.13613-13626.2004. PMID 15564472.
- ↑ 28.0 28.1 28.2 Gupta S, Takhar PP, Degenkolbe R, Koh CH, Zimmermann H, Yang CM, Guan Sim K, Hsu SI, Bernard HU (2003). "The human papillomavirus type 11 and 16 E6 proteins modulate the cell-cycle regulator and transcription cofactor TRIP-Br1" (– Scholar search). Virology 317 (1): 155–64. doi:10.1016/j.virol.2003.08.008. PMID 14675634. http://www.gis.nus.edu.sg/homepage/upload%5Cpublications%5C1012.pdf.
- ↑ 29.0 29.1 29.2 Glaunsinger BA, Lee SS, Thomas M, Banks L, Javier R (2000). "Interactions of the PDZ-protein MAGI-1 with adenovirus E4-ORF1 and high-risk papillomavirus E6 oncoproteins". Oncogene 19 (46): 5270–80. doi:10.1038/sj.onc.1203906. PMID 11077444. http://www.nature.com/onc/journal/v19/n46/full/1203906a.html.
- ↑ "iHOP information Hyperlinked over Proteins UBE3A". http://www.ihop-net.org/UniPub/iHOP/gs/92974.html. Retrieved 2007-05-01.
- ↑ "Biochemistry, Nottingham University - 3.0 Enzymes of the Ubiquitin Pathway". http://www.nottingham.ac.uk/biochemcourses/students/ub/ubenz.html. Retrieved 2007-05-01.
- ↑ Kelley ML, Keiger KE, Lee CJ, Huibregtse JM (2005). "The global transcriptional effects of the human papillomavirus E6 protein in cervical carcinoma cell lines are mediated by the E6AP ubiquitin ligase". J. Virol. 79 (6): 3737–47. doi:10.1128/JVI.79.6.3737-3747.2005. PMID 15731267. PMC 1075713. http://jvi.asm.org/cgi/content/abstract/79/6/3737.
- ↑ 33.0 33.1 Kiyono T, Hiraiwa A, Fujita M, Hayashi Y, Akiyama T, Ishibashi M (1997). "Binding of high-risk human papillomavirus E6 oncoproteins to the human homologue of the Drosophila discs large tumor suppressor protein". Proc. Natl. Acad. Sci. U.S.A. 94 (21): 11612–6. doi:10.1073/pnas.94.21.11612. PMID 9326658. PMC 23554. http://www.pnas.org/cgi/content/full/94/21/11612#abs.
- ↑ Nishimura A, Nakahara T, Ueno T, et al. (2006). "Requirement of E7 oncoprotein for viability of HeLa cells". Microbes Infect. 8 (4): 984–93. doi:10.1016/j.micinf.2005.10.015. PMID 16500131.
- ↑ Jackson ME, Pennie WD, McCaffery RE, Smith KT, Grindlay GJ, Campo MS (1991). "The B subgroup bovine papillomaviruses lack an identifiable E6 open reading frame". Mol. Carcinog. 4 (5): 382–7. doi:10.1002/mc.2940040510. PMID 1654923.
- ↑ Carter JJ, Wipf GC, Madeleine MM, Schwartz SM, Koutsky LA, Galloway DA (2006). "Identification of human papillomavirus type 16 L1 surface loops required for neutralization by human sera". J. Virol. 80 (10): 4664–72. doi:10.1128/JVI.80.10.4664-4672.2006. PMID 16641259.
- ↑ Campos SK, Ozbun MA (2009). "Two highly conserved cysteine residues in HPV16 L2 form an intramolecular disulfide bond and are critical for infectivity in human keratinocytes". PLoS ONE 4 (2): e4463. doi:10.1371/journal.pone.0004463. PMID 19214230.
- ↑ Richards RM, Lowy DR, Schiller JT, Day PM (2006). "Cleavage of the papillomavirus minor capsid protein, L2, at a furin consensus site is necessary for infection". Proc. Natl. Acad. Sci. U.S.A. 103 (5): 1522–7. doi:10.1073/pnas.0508815103. PMID 16432208.
- ↑ Yang R, Yutzy WH, Viscidi RP, Roden RB (2003). "Interaction of L2 with beta-actin directs intracellular transport of papillomavirus and infection". J. Biol. Chem. 278 (14): 12546–53. doi:10.1074/jbc.M208691200. PMID 12560332.
- ↑ Bossis I, Roden RB, Gambhira R, et al. (2005). "Interaction of tSNARE syntaxin 18 with the papillomavirus minor capsid protein mediates infection". J. Virol. 79 (11): 6723–31. doi:10.1128/JVI.79.11.6723-6731.2005. PMID 15890910.
- ↑ Pastrana DV, Gambhira R, Buck CB, et al. (2005). "Cross-neutralization of cutaneous and mucosal Papillomavirus types with anti-sera to the amino terminus of L2". Virology 337 (2): 365–72. doi:10.1016/j.virol.2005.04.011. PMID 15885736.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||