Opabinia
| Opabinia Fossil range: Middle Cambrian |
|
|---|---|
 |
|
| Artist's impression of Opabinia at the sea floor | |
| Scientific classification | |
| Domain: | Eukaryota |
| (unranked): | Opisthokonta |
| Kingdom: | Animalia |
| Subkingdom: | Eumetazoa |
| (unranked): | Bilateria Protostomia |
| Superphylum: | Ecdysozoa |
| (unranked): | Panarthropoda |
| Phylum: | Lobopodia? |
| Class: | †Dinocaridida |
| Order: | Radiodonta |
| Family: | Opabinidae |
| Genus: | Opabinia Walcott, 1912 |
| Species: | O. regalis |
| Binomial name | |
| Opabinia regalis Walcott, 1912 |
|
Opabinia is an animal genus found in Cambrian fossil deposits. Its sole species, Opabinia regalis, is known from the Middle Cambrian Burgess Shale of British Columbia. Fewer than twenty good specimens have been described; 3 specimens of Opabinia are known from the Greater Phyllopod bed, where they comprise < 0.1% of the community.[1]. Opabinia was a soft-bodied animal of modest size, and its segmented body had lobes along the sides and a fan-shaped tail. The head shows unusual features: five eyes, a mouth under the head and facing backwards, and a proboscis that probably passed food to the mouth. Opabinia probably lived on the seafloor, using the proboscis to seek out small, soft food.
When the first thorough examination of Opabinia in 1975 revealed its unusual features, it was thought to be unrelated to any known phylum, although possibly related to a hypothetical ancestor of arthropods and of annelid worms. However other finds, most notably Anomalocaris, suggested that it belonged to a group of animals that were closely related to the ancestors of arthropods and of which the living animals onychophorans and tardigrades may also be members.
In the 1970s there was an ongoing debate about whether multi-celled animals appeared suddenly during the Early Cambrian, in an event called the Cambrian explosion, or had arisen earlier but without leaving fossils. At first Opabinia was regarded as strong evidence for the "explosive" hypothesis. Later the discovery of a whole series of similar lobopod animals, some with closer resemblances to arthropods, and the development of the idea of stem groups suggested that the Early Cambrian was a time of relatively fast evolution but one that could be understood without assuming any unique evolutionary processes.
Contents |
History of discovery

Charles Doolittle Walcott found in the Burgess Shale nine almost complete fossils of Opabinia regalis and a few of what he classified as Opabinia media, and published a description of all of these in 1912. The generic name is derived from Opabin pass between Mount Hungabee and Mount Biddle, southeast of Lake O'Hara, British Columbia, Canada.[2] In 1966-1967 Harry B. Whittington found another good specimen,[3] and in 1975 he published a detailed description based on very thorough dissection of some specimens and photographs of these specimens lit from a variety of angles. Whittington's analysis did not cover Opabinia media: Walcott's specimens of this species could not be identified in his collection. In 1960 Russian paleontologists described specimens which they found in the Norilsky region of Siberia and labelled Opabinia norilica, but these fossils were poorly preserved, and Whittington did not feel they provided enough information to be classified as members of the genus Opabinia.[2]
Occurrence

All the recognized Opabinia specimens found so far come from the "Phyllopod bed" of the Burgess Shale, in the Canadian Rockies of British Columbia.[2]
In 1997 Briggs and Nedin reported from South Australia a new specimen of Myoscolex which was much better preserved than previous specimens, leading them to conclude that it was a close relative of Opabinia[4] - although this interpretation was later questioned by Dzik, who instead concluded that Myoscolex was an annelid worm.[5]
Description
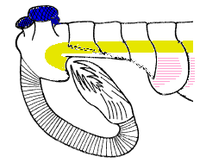
Opabinia looks so strange that the audience at the first presentation of Whittington's analysis laughed.[2] The length of Opabinia regalis from head to tail ranged between 4 centimetres (1.6 in) and 7 centimetres (2.8 in). The animal also had a hollow proboscis, whose total length was about one third of the body's and which projected down from under the head and then curved forwards and upwards. The proboscis was striated like a vacuum cleaner's hose and probably flexible, and it ended with a claw-like structure whose inner edges bore spines that projected inwards and forwards. The head bore five eyes: two on stalks near the front and fairly close to the middle of the head, pointing upwards and forwards; two larger eyes, also stalked, near the rear and outer edges of the head, pointing upwards and sideways; and a single eye with a shorter stalk between the larger pair of stalked eyes, pointing upwards. It has been assumed that the eyes were all compound, like arthropods' lateral eyes,[2] but this reconstruction, which is not backed up by any evidence,[2] is "somewhat fanciful".[6] The mouth was under the head, behind the proboscis, and pointed backwards, so that the digestive tract formed a U-bend on its way towards the rear of the animal. The proboscis appeared sufficiently long and flexible to reach the mouth.[2]
The main part of the body was typically about 5 millimetres (0.20 in) wide and had 15 segments, on each of which there was a pair of lobes (flaps) pointing downwards and outwards. The lobes overlapped so that the front of each was covered by the rear edge of the one ahead of it. The body ended with what looked like a single conical segment bearing three pairs of overlapping lobes that pointed up and out, forming a tail like a V-shaped double fan.[2]

Interpretations of other features of Opabinia fossils differ. Since the animals did not have mineralized armor nor even tough organic exoskeletons like those of insects, their bodies were flattened as they were buried and fossilized, and smaller or internal features appear as markings within the outlines of the fossils.[2][7]
Whittington interpreted as gills some markings on the upper surfaces of all lobes except the first on each side, and thought that these gills were flat underneath, had overlapping layers on top, and attached to the bases of the lobes.[2] Budd (1996) thought they consisted of separate "blades" attached along the front edges of the lobes. He also found marks inside the lobes' front edges that he interpreted as internal channels connecting the gills to the interior of the body, much as Whittington interpreted the mark along the proboscis as an internal channel.[7] On the other hand Bergström (1986) interpreted both the "gills" and the lobes as part of a covering over the upper surface of the body.[8]
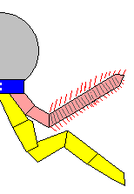
filaments
branch
Whittington found evidence of near-triangular features along the body, and concluded that they were internal structures, most likely sideways extensions of the gut.[2] Chen et al. (1994) interpreted them as contained within the lobes along the sides.[9] Budd thought the "triangles" were too wide to fit within Opabinia’s slender body, and that cross-section views showed they were attached separately from and lower than the lobes, and extended below the body. He therefore interpreted the "triangles" as short, fleshy, conical legs. He also found small mineralized patches at the tips of some, and interpreted these as claws.[7] Zhang and Briggs (2007) analyzed the chemical composition of the "triangles", and concluded that they had the same composition as the gut, and therefore agreed with Whittington that they were part of the digestive system. Instead they regarded Opabinia′s lobe+gill arrangement as an early form of the biramous limbs which are seen in trilobites and crustaceans and which may be the original form in all arthropods.[10]
Lifestyle
The way in which the Burgess Shale animals were buried, by a mudslide or a sediment-laden current that acted as a sandstorm, suggests they lived on the surface of the seafloor. Opabinia probably used its proboscis to search the sediment for food particles and pass them to its mouth. Since there is no sign of anything that might function as jaws, its food was presumably small and soft. Whittington, believing that Opabinia had no legs, thought that it crawled on its lobes and that it could also have swum slowly by flapping the lobes, especially if it timed the movements to create "Mexican waves". On the other hand he thought the body was not flexible enough to allow fish-like undulations of the whole body.[2]
Classification

Considering how paleontologists' reconstructions of Opabinia differ, it is not surprising that the animal's classification is still debated.[7] Walcott, the original describer, considered it to be an anostracan crustacean, while Leif Størmer, following earlier work by Percy Raymond, thought that it belonged to the so-called "trilobitoids". After his thorough analysis Whittington concluded that Opabinia was no arthropod, as he found no evidence for arthropodan jointed limbs, and nothing like the flexible, probably fluid-filled proboscis was known in arthropods.[2] Although he left Opabinia′s classification above the family level open, the annulated but not articulated body and the unusual lateral lobes with gills persuaded him that it may have been a representative of the ancestral stock from which both the annelids and arthropods arose.[2]
In 1985, Derek Briggs and Whittington published a description of Anomalocaris, also from the Burgess Shale. Swedish palaeontologist Jan Bergström suggested that the two animals were related, as they shared lateral flaps with gills, stalked eyes and other features; and he classified them as primitive arthropods, although he considered that arthropods are not a single phylum.[8]

|
||||||||||||||||||||||||||||
In 1996 Graham Budd found what he considered evidence of short, un-jointed legs in Opabinia,[7] His examination of the anomalocarid Kerygmachela from the Sirius Passet lagerstätte, about 518 million years ago and over 10M years older than the Burgess Shale, convinced him that this had similar legs.[11][12] He considered the legs of these two genera very similar to those of the Burgess Shale Aysheaia and the modern onychophorans, which are regarded as closely related to the ancestors of arthropods. After examining several sets of features shared by these and similar lobopods he drew up a "broad-scale reconstruction of the arthropod stem-group", in other words of arthropods and what he considered to be their evolutionary "aunts" and "cousins". One striking feature of this family tree is that modern tardigrades may be Opabinia’s closest living evolutionary relatives.[7]
Although Zhang and Briggs (2007) disagreed with Budd's diagnosis that Opabinia’s "triangles" were legs, the resemblance they saw between Opabinia’s lobe+gill arrangement and arthropods' biramous limbs led them to conclude that Opabinia was very closely related to arthropods. In fact they presented a family tree very similar to Budd's except that theirs did not mention tardigrades.[10]
Theoretical significance
Opabinia made it clear how little was known about soft-bodied animals, which do not usually leave fossils.[2] When Whittington described it in the mid-1970s, there was already a vigorous debate about the early evolution of animals. Preston Cloud argued in 1948 and 1968 that the process was "explosive",[13] and in the early 1970s Niles Eldredge and Stephen Jay Gould developed their theory of punctuated equilibrium, which views evolution as long intervals of near-stasis "punctuated" by short periods of rapid change.[14] On the other hand around the same time Wyatt Durham and Martin Glaessner both argued that the animal kingdom had a long Proterozoic history that was hidden by the lack of fossils.[15][16] Whittington (1975) concluded that Opabinia, and other taxa such as Marrella and Yohoia, cannot be accommodated in modern groups. This was one of the primary reasons why Gould in his book on the Burgess Shale, Wonderful Life, considered that Early Cambrian life was much more diverse and "experimental" than any later set of animals and that the Cambrian explosion was a truly dramatic event, possibly driven by unusual evolutionary mechanisms.[17] He regarded Opabinia as so important to understanding this phenomenon that he wanted to call his book Homage to Opabinia.[18]
However, other discoveries and analyses soon followed, revealing similar-looking animals such as Anomalocaris from the Burgess Shale and Kerygmachela from Sirius Passet.[8][11] Another Burgess Shale animal, Aysheaia, was considered very similar to modern Onychophora,[19] which are regarded as close relatives of arthropods.[20] Paleontologists defined a group called lobopods to include animals that are thought to be close relatives of arthropods but which lack jointed limbs. There is still debate about whether lobopods are monophyletic, i.e. whether they include all and only the descendants of a single common ancestor, and about whether arthropods are a sub-group of lobopods or a sister-group.[7]
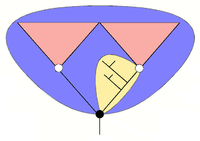
descent
While this discussion about specific fossils such as Opabinia and Anomalocaris was going on, the concept of stem groups was introduced to cover evolutionary "aunts" and "cousins". A crown group is a group of closely-related living animals plus their last common ancestor plus all its descendants. A stem group contains offshoots from members of the lineage earlier than the last common ancestor of the crown group; it is a relative concept, for example tardigrades are living animals which form a crown group in their own right, but Budd (1996) regarded them also as being a stem group relative to the arthropods.[7][21] Viewing strange-looking organisms like Opabinia in this way makes it possible to see that, while the Cambrian explosion was unusual, it can be understood in terms of normal evolutionary processes.[22]
See also
- Body plan
References
- ↑ doi:10.2110/palo.2003.P05-070R
This citation will be automatically completed in the next few minutes. You can jump the queue or expand by hand - ↑ 2.00 2.01 2.02 2.03 2.04 2.05 2.06 2.07 2.08 2.09 2.10 2.11 2.12 2.13 2.14 2.15 Whittington, H.B. (June 1975). "The enigmatic animal Opabinia regalis, Middle Cambrian Burgess Shale, British Columbia". Philosophical Transactions of the Royal Society of London: Biological Sciences 271 (910): 1–43 271. doi:10.1098/rstb.1975.0033. http://www.jstor.org/pss/2417412. Retrieved 2008-08-20. Free abstract at "The Enigmatic Animal Opabinia regalis, Middle Cambrian, Burgess Shale, British Columbia". http://adsabs.harvard.edu/abs/1975RSPTB.271....1W. Retrieved 2008-08-20.
- ↑ Gould, S.J. (1990). Wonderful Life. London: Hutchinson Radius. p. 77 and p. 189. ISBN 0091742714.
- ↑ Briggs, D.E.G., and Nedin, C. (January 1997). "The Taphonomy and Affinities of the Problematic Fossil Myoscolex from the Lower Cambrian Emu Bay Shale of South Australia". Journal of Paleontology 71 (1): 22–32. http://www.jstor.org/pss/1306537. Retrieved 2008-08-22.
- ↑ Dzik, J. (2004). "Anatomy and relationships of the Early Cambrian worm Myoscolex". Zoologica Scripta 33 (1): 57–69. doi:10.1111/j.1463-6409.2004.00136.x.
- ↑ Paulus, H.F. (2000). "Phylogeny of the Myriapoda-Crustacea-Insecta: a new attempt using photoreceptor structure*". Journal of Zoological Systematics & Evolutionary Research 38 (3): 189–208. doi:10.1046/j.1439-0469.2000.383152.x.
- ↑ 7.00 7.01 7.02 7.03 7.04 7.05 7.06 7.07 7.08 7.09 7.10 Budd, G.E. (1996). "The morphology of Opabinia regalis and the reconstruction of the arthropod stem-group". Lethaia 29 (1): 1–14. doi:10.1111/j.1502-3931.1996.tb01831.x.
- ↑ 8.0 8.1 8.2 Bergström, J. (1986). "Opabinia and Anomalocaris, unique Cambrian arthropods". Lethaia 19 (3): 241–246. doi:10.1111/j.1502-3931.1986.tb00738.x.
- ↑ Chen, J-Y., Ramsköld, L., and Gui-qing Zhou, G-Q. (May 1994). "Evidence for Monophyly and Arthropod Affinity of Cambrian Giant Predators". Science 264 (5163): 1304–1308. doi:10.1126/science.264.5163.1304. PMID 17780848. http://www.sciencemag.org/cgi/content/abstract/264/5163/1304. Retrieved 2008-08-22.
- ↑ 10.0 10.1 Zhang, X. & Briggs, D.E.G. (2007). "The nature and significance of the appendages of Opabinia from the Middle Cambrian Burgess Shale". Lethaia 40 (2): 161–173. doi:10.1111/j.1502-3931.2007.00013.x. http://www3.interscience.wiley.com/journal/117996987/abstract. Retrieved 2008-08-20.
- ↑ 11.0 11.1 Budd, G.E. (1993). "A Cambrian gilled lobopod from Greenland". Nature 364: 709–711. doi:10.1038/364709a0.
- ↑ Budd, G.E. (1997). "Stem Group Arthropods from the Lower Cambrian Sirius Passet Fauna of North Greenland". In Fortey, R.A.; Thomas, R.H.. Arthropod Relationships – Special Volume Series 55. Systematics Association.
- ↑ Cloud, P.E. (1948). "Some problems and patterns of evolution exemplified by fossil invertebrates". Evolution 2 (4): 322–350. doi:10.2307/2405523. PMID 18122310. http://jstor.org/stable/2405523. and Cloud, P.E. (1968). "Pre-metazoan evolution and the origins of the Metazoa.". In Drake, E.T.. Evolution and Environment. New Haven, Conn.: Yale University Press. pp. 1–72.
- ↑ Eldredge, N., and Gould , S.J.. "Punctuated equilibria: An alternative to phyletic gradualism.". In Schopf, T.J.M.. Models in Paleobiology. San Francisco, CA.: Freeman, Cooper & Co.. pp. 82–115.
- ↑ Bengtson, S. (2004). Early skeletal fossils. In Lipps, J.H., and Waggoner, B.M.. "Neoproterozoic- Cambrian Biological Revolutions" (PDF). Palentological Society Papers 10: 67–78. http://www.cosmonova.org/download/18.4e32c81078a8d9249800021554/Bengtson2004ESF.pdf. Retrieved 2008-07-18.
- ↑ Durham, J.W. (1971). "The fossil record and the origin of the Deuterostomata". Proceedings of the North American Paleontological Convention, Part H: 1104–1132. and Glaessner, M.F. (1972). "Precambrian palaeozoology". In Jones, J.B., and McGowran, B.. Stratigraphic Problems of the Later Precambrian and Early Cambrian. 1. University of Adelaide. pp. 43–52.
- ↑ Gould, S.J. (1989). Wonderful Life. Hutchinson Radius. pp. 124–136 and many others. ISBN 0091742714.
- ↑ Knoll, A.H. (2004). "Cambrian Redux". The First Three Billion Years of Evolution on Earth. Princeton University Press. p. 192. ISBN 9780691120294. http://books.google.com/?id=Lq0S-C6zMHAC&pg=PA192&dq=%22burgess+shale%22. Retrieved 2009-04-22.
- ↑ Robison, R.A. (January 1985). "Affinities of Aysheaia (Onychophora), with Description of a New Cambrian Species". Journal of Paleontology 59 (1): 226–235. http://www.jstor.org/pss/1304837. Retrieved 2008-08-23.
- ↑ Jacobs, D.K., Wray, C.G., Wedeen, C.J.Kostriken, R., DeSalle, R., Staton, J.L., Gates, R.D., Lindberg, D.R. (2000). "Molluscan engrailed Expression, Serial Organization, and Shell Evolution" (PDF). Evolution and Development 2 (6): 340–347. doi:10.1046/j.1525-142x.2000.00077.x. PMID 11256378. http://www.biol.sc.edu/~coull_lab/staton/ref/engrailed.pdf. Retrieved 2008-08-23.
- ↑ 21.0 21.1 Craske, A.J., and Jefferies, R.P.S. (1989). "A new mitrate from the Upper Ordovician of Norway, and a new approach to subdividing a plesion". Palaeontology 32: 69–99.
- ↑ Budd, G.E. (2003). "The Cambrian Fossil Record and the Origin of the Phyla" (Free full text). Integrative and Comparative Biology 43 (1): 157–165. doi:10.1093/icb/43.1.157. http://intl-icb.oxfordjournals.org/cgi/content/abstract/43/1/157. Retrieved 2008-08-23.
Further reading
- Bergström, J. (1987). "The Cambrian Opabinia and Anomalocaris". Lethaia 20 (2): 187–188. doi:10.1111/j.1502-3931.1987.tb00777.x.
- Briggs, D.E.G. & Whittington, H.B. (1987). "The affinities of the Cambrian animals Anomalocaris and Opabinia". Lethaia 20 (2): 185–186. doi:10.1111/j.1502-3931.1987.tb00776.x.