Mitosis

Mitosis is the process by which a eukaryotic cell separates the chromosomes in its cell nucleus into two identical sets in two nuclei.[1] It is generally followed immediately by cytokinesis, which divides the nuclei, cytoplasm, organelles and cell membrane into two cells containing roughly equal shares of these cellular components. Mitosis and cytokinesis together define the mitotic (M) phase of the cell cycle - the division of the mother cell into two daughter cells, genetically identical to each other and to their parent cell. This accounts for approximately 10% of the cell cycle.
Mitosis occurs exclusively in eukaryotic cells, but occurs in different ways in different species. For example, animals undergo an "open" mitosis, where the nuclear envelope breaks down before the chromosomes separate, while fungi such as Aspergillus nidulans and Saccharomyces cerevisiae (yeast) undergo a "closed" mitosis, where chromosomes divide within an intact cell nucleus.[2] Prokaryotic cells, which lack a nucleus, divide by a process called binary fission.
The process of mitosis is complex and highly regulated. The sequence of events is divided into phases, corresponding to the completion of one set of activities and the start of the next. These stages are interphase, prophase, prometaphase, metaphase, anaphase and telophase. During the process of mitosis the pairs of chromosomes condense and attach to fibers that pull the sister chromatids to opposite sides of the cell. The cell then divides in cytokinesis, to produce two identical daughter cells.[3]
Because cytokinesis usually occurs in conjunction with mitosis, "mitosis" is often used interchangeably with "mitotic phase". However, there are many cells where mitosis and cytokinesis occur separately, forming single cells with multiple nuclei. This occurs most notably among the fungi and slime moulds, but is found in various different groups. Even in animals, cytokinesis and mitosis may occur independently, for instance during certain stages of fruit fly embryonic development.[4] Errors in mitosis can either kill a cell through apoptosis or cause mutations that may lead to cancer.
Contents |
Overview
The primary result of mitosis is the division of the parent cell's genome into two daughter cells. The genome is composed of a number of chromosomes - complexes of tightly-coiled DNA that contain genetic information vital for proper cell function. Because each resultant daughter cell should be genetically identical to the parent cell, the parent cell must make a copy of each chromosome before mitosis. This occurs during S phase, in interphase, the period that precedes the mitotic phase in the cell cycle where preparation for mitosis occurs.[5]
Each new chromosome now contains two identical copies of itself, called sister chromatids, attached together in a specialized region of the chromosome known as the centromere. Each sister chromatid is not considered a chromosome in itself, and a chromosome always contains two sister chromatids.
In most eukaryotes, the nuclear envelope that separates the DNA from the cytoplasm disassembles. The chromosomes align themselves in a line spanning the cell. Microtubules, essentially miniature strings, splay out from opposite ends of the cell and shorten, pulling apart the sister chromatids of each chromosome.[6] As a matter of convention, each sister chromatid is now considered a chromosome, so they are renamed to sister chromosomes. As the cell elongates, corresponding sister chromosomes are pulled toward opposite ends. A new nuclear envelope forms around the separated sister chromosomes.
As mitosis completes cytokinesis is well underway. In animal cells, the cell pinches inward where the imaginary line used to be (the pinching of the cell membrane to form the two daughter cells is called the cleavage furrow), separating the two developing nuclei. In plant cells, the daughter cells will construct a new dividing cell wall between each other. Eventually, the mother cell will be split in half, giving rise to two daughter cells, each with an equivalent and complete copy of the original genome.
Prokaryotic cells undergo a process similar to mitosis called binary fission. However, prokaryotes cannot be properly said to undergo mitosis because they lack a nucleus and only have a single chromosome with no centromere.[7]
Phases of cell cycle and mitosis
Interphase
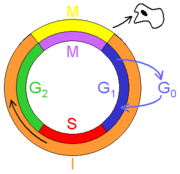
The mitotic phase is a relatively short period of the cell cycle. It alternates with the much longer interphase, where the cell prepares itself for cell division. Interphase is therefore not part of mitosis. Interphase is divided into three phases, G1 (first gap), S (synthesis), and G2 (second gap). During all three phases, the cell grows by producing proteins and cytoplasmic organelles. However, chromosomes are replicated only during the S phase. Thus, a cell grows (G1), continues to grow as it duplicates its chromosomes (S), grows more and prepares for mitosis (G2), and finally divides (M) before restarting the cycle.[5]
Preprophase
In plant cells only, prophase is preceded by a pre-prophase stage. In highly vacuolated plant cells, the nucleus has to migrate into the center of the cell before mitosis can begin. This is achieved through the formation of a phragmosome, a transverse sheet of cytoplasm that bisects the cell along the future plane of cell division. In addition to phragmosome formation, preprophase is characterized by the formation of a ring of microtubules and actin filaments (called preprophase band) underneath the plasma membrane around the equatorial plane of the future mitotic spindle. This band marks the position where the cell will eventually divide. The cells of higher plants (such as the flowering plants) lack centrioles: with microtubules forming a spindle on the surface of the nucleus and then being organized into a spindle by the chromosomes themselves, after the nuclear membrane breaks down.[8] The preprophase band disappears during nuclear envelope disassembly and spindle formation in prometaphase.[9]
|




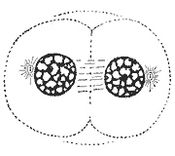
Prophase

Normally, the genetic material in the nucleus is in a loosely bundled coil called chromatin. At the onset of prophase, chromatin condenses together into a highly ordered structure called a chromosome. Since the genetic material has already been duplicated earlier in S phase, the replicated chromosomes have two sister chromatids, bound together at the centromere by the cohesion complex. Chromosomes are visible at high magnification through a light microscope.
Close to the nucleus are structures called centrosomes, which are made of a pair of centrioles. The centrosome is the coordinating center for the cell's microtubules. A cell inherits a single centrosome at cell division, which replicates before a new mitosis begins, giving a pair of centrosomes. The two centrosomes nucleate microtubules (which may be thought of as cellular ropes or poles) to form the spindle by polymerizing soluble tubulin. Molecular motor proteins then push the centrosomes along these microtubules to opposite sides of the cell. Although centrioles help organize microtubule assembly, they are not essential for the formation of the spindle, since they are absent from plants,[8] and centrosomes are not always used in meiosis.[10]
Prometaphase
The nuclear envelope disassembles and microtubules invade the nuclear space. This is called open mitosis, and it occurs in most multicellular organisms. Fungi and some protists, such as algae or trichomonads, undergo a variation called closed mitosis where the spindle forms inside the nucleus or its microtubules are able to penetrate an intact nuclear envelope.[11][12]
Each chromosome forms two kinetochores at the centromere, one attached at each chromatid. A kinetochore is a complex protein structure that is analogous to a ring for the microtubule hook; it is the point where microtubules attach themselves to the chromosome.[13] Although the kinetochore structure and function are not fully understood, it is known that it contains some form of molecular motor.[14] When a microtubule connects with the kinetochore, the motor activates, using energy from ATP to "crawl" up the tube toward the originating centrosome. This motor activity, coupled with polymerisation and depolymerisation of microtubules, provides the pulling force necessary to later separate the chromosome's two chromatids.[14]
When the spindle grows to sufficient length, kinetochore microtubules begin searching for kinetochores to attach to. A number of nonkinetochore microtubules find and interact with corresponding nonkinetochore microtubules from the opposite centrosome to form the mitotic spindle.[15] Prometaphase is sometimes considered part of prophase.
In the fishing pole analogy, the kinetochore would be the "hook" that catches a sister chromatid or "fish". The centrosome acts as the "reel" that draws in the spindle fibers or "fishing line".
Metaphase

As microtubules find and attach to kinetochores in prometaphase, the centromeres of the chromosomes convene along the metaphase plate or equatorial plane, an imaginary line that is equidistant from the two centrosome poles.[15] This even alignment is due to the counterbalance of the pulling powers generated by the opposing kinetochores, analogous to a tug-of-war between people of equal strength. In certain types of cells, chromosomes do not line up at the metaphase plate and instead move back and forth between the poles randomly, only roughly lining up along the midline. Metaphase comes from the Greek μετα meaning "after."
Because proper chromosome separation requires that every kinetochore be attached to a bundle of microtubules (spindle fibres), it is thought that unattached kinetochores generate a signal to prevent premature progression to anaphase without all chromosomes being aligned. The signal creates the mitotic spindle checkpoint.[16]
Anaphase
When every kinetochore is attached to a cluster of microtubules and the chromosomes have lined up along the metaphase plate, the cell proceeds to anaphase (from the Greek ανα meaning “up,” “against,” “back,” or “re-”).
Two events then occur; First, the proteins that bind sister chromatids together are cleaved, allowing them to separate. These sister chromatids, which have now become distinct sister chromosomes, are pulled apart by shortening kinetochore microtubules and move toward the respective centrosomes to which they are attached. Next, the nonkinetochore microtubules elongate, pulling the centrosomes (and the set of chromosomes to which they are attached) apart to opposite ends of the cell. The force that causes the centrosomes to move towards the ends of the cell is still unknown, although there is a theory that suggests that the rapid assembly and breakdown of microtubules may cause this movement.[17]
These two stages are sometimes called early and late anaphase. Early anaphase is usually defined as the separation of the sister chromatids, while late anaphase is the elongation of the microtubules and the chromosomes being pulled farther apart. At the end of anaphase, the cell has succeeded in separating identical copies of the genetic material into two distinct populations.
Telophase
Telophase (from the Greek τελος meaning "end") is a reversal of prophase and prometaphase events. It "cleans up" the after effects of mitosis. At telophase, the nonkinetochore microtubules continue to lengthen, elongating the cell even more. Corresponding sister chromosomes attach at opposite ends of the cell. A new nuclear envelope, using fragments of the parent cell's nuclear membrane, forms around each set of separated sister chromosomes. Both sets of chromosomes, now surrounded by new nuclei, unfold back into chromatin. Mitosis is complete, but cell division is not yet complete.
Cytokinesis
Cytokinesis is often mistakenly thought to be the final part of telophase; however, cytokinesis is a separate process that begins at the same time as telophase. Cytokinesis is technically not even a phase of mitosis, but rather a separate process, necessary for completing cell division. In animal cells, a cleavage furrow (pinch) containing a contractile ring develops where the metaphase plate used to be, pinching off the separated nuclei.[18] In both animal and plant cells, cell division is also driven by vesicles derived from the Golgi apparatus, which move along microtubules to the middle of the cell.[19] In plants this structure coalesces into a cell plate at the center of the phragmoplast and develops into a cell wall, separating the two nuclei. The phragmoplast is a microtubule structure typical for higher plants, whereas some green algae use a phycoplast microtubule array during cytokinesis.[20] Each daughter cell has a complete copy of the genome of its parent cell. The end of cytokinesis marks the end of the M-phase.
Significance
Mitosis is important for the maintenance of the chromosomal set; each cell formed receives chromosomes that are alike in composition and equal in number to the chromosomes of the parent cell. Transcription is generally believed to cease during mitosis, but epigenetic mechanisms such as bookmarking function during this stage of the cell cycle to ensure that the "memory" of which genes were active prior to entry into mitosis are transmitted to the daughter cells.[21]
Consequences of errors

Although errors in mitosis are rare, the process may go wrong, especially during early cellular divisions in the zygote. Mitotic errors can be especially dangerous to the organism because future offspring from this parent cell will carry the same disorder.
In non-disjunction, a chromosome may fail to separate during anaphase. One daughter cell will receive both sister chromosomes and the other will receive none. This results in the former cell having three chromosomes containing the same genes (two sisters and a homologue), a condition known as trisomy, and the latter cell having only one chromosome (the homologous chromosome), a condition known as monosomy. These cells are considered aneuploid, a condition often associated with cancer.[22]
Mitosis is a traumatic process. The cell goes through dramatic changes in ultrastructure, its organelles disintegrate and reform in a matter of hours, and chromosomes are jostled constantly by probing microtubules. Occasionally, chromosomes may become damaged. An arm of the chromosome may be broken and the fragment lost, causing deletion. The fragment may incorrectly reattach to another, non-homologous chromosome, causing translocation. It may reattach to the original chromosome, but in reverse orientation, causing inversion. Or, it may be treated erroneously as a separate chromosome, causing chromosomal duplication. The effect of these genetic abnormalities depends on the specific nature of the error. It may range from no noticeable effect to cancer induction or organism death.
Endomitosis
Endomitosis is a variant of mitosis without nuclear or cellular division, resulting in cells with many copies of the same chromosome occupying a single nucleus. This process may also be referred to as endoreduplication and the cells as endoploid.[4] An example of a cell that goes through endomitosis is the megakaryocyte.[23]
Timeline in pictures
Real mitotic cells can be visualized through the microscope by staining them with fluorescent antibodies and dyes. These light micrographs are included below.
 Early prophase: Nonkinetochore microtubules, shown as green strands, have established a matrix around the degrading nucleus, in blue. The green nodules are the centrosomes. |
 Late metaphase: The centrosomes have moved to the poles of the cell and have established the mitotic spindle. The chromosomes, in light blue, have all assembled at the metaphase plate, except for one. |
 Anaphase: Lengthening nonkinetochore microtubules push the two sets of chromosomes further apart. |
See also
- Meiosis
- Cytoskeleton
- Motor protein
- Aneuploidy
- Chromosomal disorder
- Binary fission
References
- ↑ Rubenstein, Irwin, and Susan M. Wick. "Cell." World Book Online Reference Center. 2008. 12 January 2008 <http://www.worldbookonline.com/wb/Article?id=ar102240>
- ↑ De Souza CP, Osmani SA (2007). "Mitosis, not just open or closed". Eukaryotic Cell 6 (9): 1521–7. doi:10.1128/EC.00178-07. PMID 17660363.
- ↑ Maton, Anthea; Hopkins, Jean Johnson, Susan LaHart, David, Quon Warner, David, Wright, Jill D (1997). Cells: Building Blocks of Life. New Jersey: Prentice Hall. pp. 70–4. ISBN 0-13423476-6.
- ↑ 4.0 4.1 Lilly M, Duronio R (2005). "New insights into cell cycle control from the Drosophila endocycle". Oncogene 24 (17): 2765–75. doi:10.1038/sj.onc.1208610. PMID 15838513.
- ↑ 5.0 5.1 Blow J, Tanaka T (2005). "The chromosome cycle: coordinating replication and segregation. Second in the cycles review series". EMBO Rep 6 (11): 1028–34. doi:10.1038/sj.embor.7400557. PMID 16264427.
- ↑ Zhou J, Yao J, Joshi H (2002). "Attachment and tension in the spindle assembly checkpoint". J Cell Sci 115 (Pt 18): 3547–55. doi:10.1242/jcs.00029. PMID 12186941.
- ↑ Nanninga N (2001). "Cytokinesis in prokaryotes and eukaryotes: common principles and different solutions". Microbiol Mol Biol Rev 65 (2): 319–33. doi:10.1128/MMBR.65.2.319-333.2001. PMID 11381104.
- ↑ 8.0 8.1 Lloyd C, Chan J (February 2006). "Not so divided: the common basis of plant and animal cell division". Nature reviews. Molecular cell biology 7 (2): 147–52. doi:10.1038/nrm1831. PMID 16493420.
- ↑ Raven, Peter H.; Ray F. Evert, Susan E. Eichhorn (2005). Biology of Plants (7th ed.). New York: W.H. Freeman and Company Publishers. pp. 58–67. ISBN 0-7167-1007-2.
- ↑ Varmark H (April 2004). "Functional role of centrosomes in spindle assembly and organization". Journal of cellular biochemistry 91 (5): 904–14. doi:10.1002/jcb.20013. PMID 15034926.
- ↑ Heywood P. (1978). "Ultrastructure of mitosis in the chloromonadophycean alga Vacuolaria virescens". J Cell Sci. 31: 37–51. PMID 670329.
- ↑ Ribeiro K, Pereira-Neves A, Benchimol M (2002). "The mitotic spindle and associated membranes in the closed mitosis of trichomonads". Biol Cell 94 (3): 157–72. doi:10.1016/S0248-4900(02)01191-7. PMID 12206655.
- ↑ Chan G, Liu S, Yen T (2005). "Kinetochore structure and function". Trends Cell Biol 15 (11): 589–98. doi:10.1016/j.tcb.2005.09.010. PMID 16214339.
- ↑ 14.0 14.1 Maiato H, DeLuca J, Salmon E, Earnshaw W (2004). "The dynamic kinetochore-microtubule interface". J Cell Sci 117 (Pt 23): 5461–77. doi:10.1242/jcs.01536. PMID 15509863.
- ↑ 15.0 15.1 Winey M, Mamay C, O'Toole E, Mastronarde D, Giddings T, McDonald K, McIntosh J (1995). "Three-dimensional ultrastructural analysis of the Saccharomyces cerevisiae mitotic spindle". J Cell Biol 129 (6): 1601–15. doi:10.1083/jcb.129.6.1601. PMID 7790357.
- ↑ Chan G, Yen T (2003). "The mitotic checkpoint: a signaling pathway that allows a single unattached kinetochore to inhibit mitotic exit". Prog Cell Cycle Res 5: 431–9. PMID 14593737.
- ↑ Kenneth R. Miller. (2000). "Anaphase". Biology.: 169–170.
- ↑ Glotzer M (2005). "The molecular requirements for cytokinesis". Science 307 (5716): 1735–9. doi:10.1126/science.1096896. PMID 15774750.
- ↑ Albertson R, Riggs B, Sullivan W (2005). "Membrane traffic: a driving force in cytokinesis". Trends Cell Biol 15 (2): 92–101. doi:10.1016/j.tcb.2004.12.008. PMID 15695096.
- ↑ Raven, Peter H.; Ray F. Evert, Susan E. Eichhorn (2005). Biology of Plants (7th ed.). New York: W.H. Freeman and Company Publishers. pp. 64–7, 328–9. ISBN 0-7167-1007-2.
- ↑ Zhou G, Liu D, Liang C (2005). "Memory mechanisms of active transcription during cell division". Bioessays 27 (12): 1239–45. doi:10.1002/bies.20327. PMID 16299763.
- ↑ Draviam V, Xie S, Sorger P (2004). "Chromosome segregation and genomic stability". Curr Opin Genet Dev 14 (2): 120–5. doi:10.1016/j.gde.2004.02.007. PMID 15196457.
- ↑ Italiano JE, Shivdasani RA (2003). "Megakaryocytes and beyond: the birth of platelets". J. Thromb. Haemost. 1 (6): 1174–82. doi:10.1046/j.1538-7836.2003.00290.x. PMID 12871316.
Further reading
- Morgan, David L. (2007). The cell cycle: principles of control. London: Published by New Science Press in association with Oxford University Press. ISBN 0-9539181-2-2.
- Alberts B, Johnson A, Lewis J, Raff M, Roberts K, and Walter P (2002). "Mitosis". Molecular Biology of the Cell. Garland Science. http://www.ncbi.nlm.nih.gov/books/bv.fcgi?highlight=mitosis&rid=mboc4.section.3349. Retrieved 2006-01-22.
- Campbell, N. and Reece, J. (December 2001). "The Cell Cycle". Biology (6th ed.). San Francisco: Benjamin Cummings/Addison-Wesley. pp. 217–224. ISBN 0-8053-6624-5.
- Cooper, G. (2000). "The Events of M Phase". The Cell: A Molecular Approach. Sinaeur Associates, Inc. http://www.ncbi.nlm.nih.gov/books/bv.fcgi?highlight=M%20Phase,Events&rid=cooper.section.2470. Retrieved 2006-01-22.
- Freeman, S (2002). "Cell Division". Biological Science. Upper Saddle River, NJ: Prentice Hall. pp. 155–174. ISBN 0-13-081923-9.
- Lodish H, Berk A, Zipursky L, Matsudaira P, Baltimore D, Darnell J (2000). "Overview of the Cell Cycle and Its Control". Molecular Cell Biology. W.H. Freeman. http://www.ncbi.nlm.nih.gov/books/bv.fcgi?highlight=Overview,Control,Cell+Cycle&rid=mcb.section.3463. Retrieved 2006-01-22.
External links
- Mitosis Animation.
- Mitosis (Flash Animation)
- Video of a live amphibian lung cell undergoing mitosis.
- A Flash animation comparing Mitosis and Meiosis
- Animation from the Life Sciences Dept of University of North Texas
- Studying Mitosis in Cultured Mammalian Cells
- General K-12 classroom resources for Mitosis
- CCO The Cell-Cycle Ontology
|
|||||||||||||||||||||||||||||||