Fertilisation
- For soil improvement see Fertiliser.
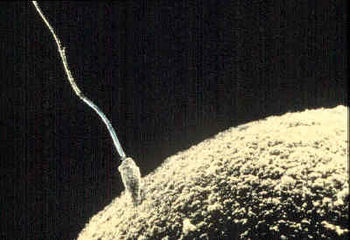
Fertilisation (also known as conception, fecundation and syngamy), is the fusion of gametes to produce a new organism. In animals, the process involves the fusion of an ovum with a sperm, which eventually leads to the development of an embryo. Depending on the animal species, the process can occur within the body of the female in internal fertilisation, or outside in the case of external fertilisation
The entire process of development of new individuals is called procreation, the act of species reproduction.
Contents |
Fertilisation in plants
Flowering plants
After the carpel is pollinated, the pollen grain germinates in a response to a sugary fluid secreted by the mature stigma (mainly sucrose). From each pollen grain, a pollen tube grows out that attempts to travel to the ovary by creating a path through the female tissue. The vegetative (or tube) and generative nuclei of the pollen grain pass into its respective pollen tube. After the pollen grain adheres to the stigma of the carpel (female reproductive structure) a pollen tube grows and penetrates the ovule through a tiny pore called a micropyle.
The pollen tube does not directly reach the ovary in a straight line. It travels near the skin of the style and curls to the bottom of the ovary, then near the receptacle, it breaks through the ovule through the micropyle (an opening in the ovule wall) and the pollen tube "bursts" into the embryo sac. After being fertilised, the ovary starts to swell and will develop into the fruit.[1] With multi-seeded fruits, multiple grains of pollen are necessary for syngamy with each ovule. The growth of the pollen tube is controlled by the vegetative (or tube) cytoplasm. Hydrolytic enzymes are secreted by the pollen tube that digest the female tissue as the tube grows down the stigma and style; the digested tissue is used as a nutrient source for the pollen tube as it grows. During pollen tube growth toward the ovary, the generative nucleus divides to produce two separate sperm nuclei (haploid number of chromosomes)[2] - a growing pollen tube therefore contains three separate nuclei, two sperm and one tube.[3] The sperms are interconnected and dimorphic, the large one, in a number of plants, is also linked to the tube nucleus and the interconnected sperm and tube nucleuses form the "male germ unit".[4]
Double fertilisation is the process in angiosperms (flowering plants) in which two sperm nuclei from each pollen tube fertilise two cells in an ovary. After the pollen tube reaches the ovary the pollen tube nucleus disintegrates and the two sperm cells are released into the ovary; one of the two sperm cells fertilises the egg cell (at the bottom of the ovule near the micropyle), forming a diploid (2n) zygote. This is the point when fertilisation actually occurs. Note that pollination and fertilisation are two separate processes. The other sperm cell fuses with two haploid polar nuclei (contained in the central cell) in the centre of the embryo sac (or ovule). The resulting cell is triploid (3n). This triploid cell divides through mitosis and forms the endosperm, a nutrient-rich tissue, inside the seed.
The two central cell maternal nuclei (polar nuclei) that contribute to the endosperm arise by mitosis from a single meiotic product. Therefore, maternal contribution to the genetic constitution of the triploid endosperm is different from that of the embryo.
Double fertilisation occurs only in angiosperm plants. One primitive species of flowering plant, Nuphar polysepala, has endosperm that is diploid, resulting from the fusion of a pollen nucleus with one, rather than two, maternal nuclei. It is believed that early in the development of angiosperm linages, there was a duplication in this mode of reproduction, producing seven-celled/eight-nucleate female gametophytes, and triploid endosperms with a 2:1 maternal to paternal genome ratio.[5]
The process is easy to visualise if one looks at maize silk, which is the female flower of corn. Pollen from the tassel (the male flower) falls on the sticky external portion of the silk, and then pollen tubes grow down the silk to the attached ovule. The dried silk remains inside the husk of the ear as the seeds mature; if one carefully removes the husk, the floral structures may be seen.
In many plants, the development of the flesh of the fruit is proportional to the percentage of fertilised ovules. For example, with watermelon, about a thousand grains of pollen must be delivered and spread evenly on the three lobes of the stigma to make a normal sized and shaped fruit.
Fertilisation in animals
The mechanics behind fertilisation has been studied extensively in sea urchins and mice. This research addresses the question of how the sperm and the appropriate egg find each other and the question of how only one sperm gets into the egg and delivers its contents. There are three steps to fertilisation that ensure species-specificity:
- Chemotaxis
- Sperm activation/acrosomal reaction
- Sperm/egg adhesion.
Internal vs. External
Consideration as to whether an animal (more specifically a vertebrate) uses internal or external fertilisation is often dependent on the method of birth. Oviparous animals laying eggs with thick calcium shells, such as chickens, or thick leathery shells generally reproduce via internal fertilisation so that the sperm fertilise the egg without having to pass through the thick, protective, tertiary layer of the egg. Ovoviviparous and euviviparous animals also use internal fertilisation. It is important to note that although some organisms reproduce via amplexus, they may still use internal fertilisation, as with some salamanders. Advantages to internal fertilisation include: minimal waste of gametes; greater chance of individual egg fertilisation, relatively "longer" time period of egg protection, and selective fertilisation; many females have the ability to store sperm for extended periods of time and can fertilise their eggs at their own desire.
Oviparous animals producing eggs with thin tertiary membranes or no membranes at all, on the other hand, use external fertilisation methods. Advantages to external fertilisation include: minimal contact and transmission of bodily fluids; decreasing the risk of disease transmission, and greater genetic variation (especially during broadcast spawning external fertilisation methods).
Sea urchins

Chemotaxis was discovered as the method by which sperm find the eggs. This chemotaxis is an example of a ligand/receptor interaction. Resact is a 14 amino acid peptide purified from the jelly coat of A. punctulata that attracts the migration of sperm.
After finding the egg, the sperm gets through the jelly coat through a process called sperm activation. In another ligand/receptor interaction, an oligosaccharide component of the egg binds and activates a receptor on the sperm and causes the acrosomal reaction. The acrosomal vesicles of the sperm fuse with the plasma membrane and are released. In this process, molecules bound to the acrosomal vesicle membrane, such as bindin, are exposed on the surface of the sperm. These contents digest the jelly coat and eventually the vitelline membrane. In addition to the release of acrosomal vesicles, there is explosive polymerization of actin to form a thin spike at the head of the sperm called the acrosomal process.
The sperm binds to the egg through another ligand reaction between receptors on the vitelline membrane. The sperm surface protein bindin, binds to a receptor on the vitelline membrane identified as ERB1.
Fusion of the plasma membranes of the sperm and egg are likely mediated by bindin. At the site of contact, fusion causes the formation of a fertilisation cone.
Mammals
Usually mammals rely on internal fertilisation through copulation. After a male ejaculates, a large number of sperm cells move to the upper vagina (via contractions from the vagina) through the cervix and across the length of the uterus toward the ovum. The capacitated spermatozoon and the oocyte meet and interact in the ampulla of the fallopian tube. It is probable that chemotaxis is involved in guiding the sperm to the egg, but the mechanism has yet to be worked out. However, demonstration of formyl peptide receptors (60.000 receptor/cell; higher binding capacity in the tail region) in the surface membrane of human sperms strongly supports, that - besides specific chemoattractant substances i.e. resact - professional chemoattractant ligands like formyl Met-Leu-Phe (fMLF) have also the ability to induce migration of sperm.[6]
The zona pellucida of the egg binds with the sperm. In contrast to sea urchins, the sperm binds to the egg before the acrosmal reaction. The zona pellucida is a thick layer of extracellular matrix that surrounds the egg and is similar to the role of the vitelline membrane in sea urchins. A glycoprotein in the zona pellucida, ZP3 was discovered to be responsible for egg/sperm adhesion in mice. The receptor galactosyltransferase (GalT) binds to the N-acetylglucosamine residues on the ZP3 and is important for binding with the sperm and activating the acrosome reaction. ZP3 is sufficient for sperm/egg binding but not necessary. There are two additional sperm receptors: a 250kD protein that binds to an oviduct secreted protein and SED1 which binds independently to the zona. After the acrosome reaction, it is believed that the sperm remains bound to the zona pellucida through exposed ZP2 receptors. These receptors are unknown in mice but have been identified in guinea pigs.
In mammals, binding of the spermatozoon to the GalT initiates the acrosome reaction. This process releases the enzyme hyaluronidase, which digests the matrix of hyaluronic acid in the vestments surrounding the oocyte. Fusion between the oocyte plasma membranes and sperm follows, allowing the entry of the sperm nucleus, centriole and flagellum, but not the mitochondria, into the oocyte. The fusion is likely mediated by the protein CD9 in mice (the binding homolog). The egg "activates" itself upon fusing with a single sperm cell, thereby changing its cell membrane to prevent fusion with other sperm.
This process ultimately leads to the formation of a diploid cell called a zygote. The zygote begins to divide and form a blastocyst and when it reaches the uterus, it performs implantation in the endometrium. At this point the female's pregnancy has begun. If the embryo implants in any tissue other than the uterine wall, an ectopic pregnancy results, which can be fatal to the mother.
In some animals (e.g. rabbits) the act of coitus induces ovulation by stimulating release of the pituitary hormone gonadotropin. This greatly increases the probability that coitus will result in pregnancy.
Humans
The term conception commonly refers to fertilisation, the successful fusion of gametes to form a new organism. 'Conception' is used by some to refer to implantation and is thus a subject of semantic arguments about the beginning of pregnancy, typically in the context of the abortion debate. Gastrulation, which occurs around 16 days after fertilisation, is the point in development when the implanted blastocyst develops three germ layers, the endoderm, the ectoderm and the mesoderm. It is at this point that the genetic code of the father becomes fully involved in the development of the embryo. Until this point in development, twinning is possible. Additionally, interspecies hybrids survive only until gastrulation, and have no chance of development afterward. However this stance is not entirely accepted as some human developmental biology literature refers to the "conceptus" and such medical literature refers to the "products of conception" as the post-implantation embryo and its surrounding membranes.[7] The term "conception" is not usually used in scientific literature because of its variable definition and connotation.
Fertilisation and genetic recombination
Meiosis results in a random segregation of the genes contributed from each parent. Each parent organism generally has the same genetic make-up, but differs for a fraction of their genes. Therefore, each gamete produced by a person will be genetically different from the others from that person, as well as from the gametes produced by another person. When gametes first fuse at fertilisation, the chromosomes donated by the parents are combined, and, in humans, this means that (2²²)² = 17.6x1012 chromosomally different zygotes are possible for the non-sex chromosomes, even assuming no chromosomal crossover. If crossover occurs once, then on average (4²²)² = 309x1024 genetically different zygotes are possible for every couple, not considering that crossover events can take place at most points along each chromosome. The X and Y chromosomes do not undergo crossover events, so are excluded from the calculation. Note that the mitochondrial DNA is only inherited from the maternal parent.
Parthenogenesis
Another method of fertilisation occurs among animals that normally reproduce sexually, through parthenogenesis: when the gamete of a female is not fertilised by a male, yet produces viable and unique offspring that are not clones. Only DNA from the mother is inherited, but it is not identical to her. Normal eggs of the mother become fertilised, without sperm, and development proceeds normally. This occurs naturally in several species and may be induced in others through a chemical or electrical stimulus. In 2004, Japanese researchers led by Tomohiro Kono succeeded after 457 attempts to merge the ova of two mice, the result of which developed normally into a mouse. This was achieved by blocking certain proteins that would normally prevent the possibility.[8]
See also
- Kaguya (mouse)
- Female sperm
- In vitro fertilisation
- Fetal development
- Superfetation
- Superfecundation
- Conception Cap
- Conception Device
- Proembryo
References
- ↑ Johnstone, Adam. Biology: facts & practice for A level. Oxford University Press. pp. 95. ISBN 0-19-914766-3.
- ↑ Handbook of plant science. Chichester, West Sussex, England: John Wiley. 2007. pp. 466. ISBN 978-0-470-05723-0.
- ↑ Kirk, David; Starr, Cecie (1975). Biology today. [Del Mar, Calif.]: CRM. pp. 93. ISBN 978-0-394-31093-0.
- ↑ Raghavan, V. (Valayamghat) (2006). Double fertilization : embryo and endosperm development in flowering plant. Berlin: Springer-Verlag. pp. 12. ISBN 978-3-540-27791-0.
- ↑ Friedman WE, Williams JH (February 2003). "Modularity of the angiosperm female gametophyte and its bearing on the early evolution of endosperm in flowering plants". Evolution; International Journal of Organic Evolution 57 (2): 216–30. doi:10.1111/j.0014-3820.2003.tb00257.x. PMID 12683519.
- ↑ Gnessi L, Fabbri A, Silvestroni L, Moretti C, Fraioli F, Pert CB, Isidori A. (1986). "Evidence for the presence of specific receptors for N-formyl chemotactic peptides on human spermatozoa.". J Clin Endocrinol Metab 63 (4): 841–6. doi:10.1210/jcem-63-4-841. PMID 3018025.
- ↑ Moore, K. L. & T. V. M. Persaud (2003). The Developing Human: Clinically Oriented Embryology. W. B. Saunders Company. ISBN 0-7216-6974-3.
- ↑ Kono T, Obata Y, Wu Q, et al. (April 2004). "Birth of parthenogenetic mice that can develop to adulthood". Nature 428 (6985): 860–4. doi:10.1038/nature02402. PMID 15103378. Lay summary – National Geographic (2004-04-21).
|
||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||