Sauropoda
| Sauropods Fossil range: Early Jurassic–Late Cretaceous, 188–66 Ma |
|
|---|---|
 |
|
| Mounted skeleton cast of Diplodocus carnegii | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Reptilia |
| Superorder: | Dinosauria |
| Order: | Saurischia |
| Suborder: | †Sauropodomorpha |
| Infraorder: | †Sauropoda Marsh, 1878 |
| Families | |
|
See text. |
|
Sauropoda (pronounced /sɔːˈrɒpɵdə/), or the sauropods (/ˈsɔrəpɒd/), are an infraorder of saurischian ("lizard-hipped") dinosaurs. They are notable for the enormous sizes attained by some species, and the group includes the largest animals to have ever lived on land. Well-known genera include Brachiosaurus, Diplodocus, and Apatosaurus (which, under current classification, is a senior synonym of Brontosaurus). Sauropods first appeared in the late Triassic Period, where they somewhat resembled the closely related (and possibly ancestral) group Prosauropoda. By the Late Jurassic (150 million years ago), sauropods were widespread (especially the diplodocids and brachiosaurids). By the Late Cretaceous, those groups had mainly been replaced by the titanosaurs, which had a near-global distribution. However, as with all other non-avian dinosaurs, the titanosaurs died out in the Cretaceous–Tertiary extinction event. Fossilised remains of sauropods have been found on every continent, including Antarctica.
The name Sauropoda was coined by O.C. Marsh in 1878, and is derived from the Greek for "lizard foot".[1]
Complete sauropod fossil finds are rare. Many species, especially the largest, are known only from isolated and disarticulated bones. Many near-complete specimens lack heads, tail tips and limbs.
Contents |
Description
Sauropods were herbivorous (plant-eating), usually quite long-necked quadrupeds (four-legged), often with spatulate (spatula-shaped: broad at the base, narrow at the neck) teeth. They had tiny heads, massive bodies, and tended to have long tails. Their hind legs were thick, straight, and powerful, ending in club-like feet with five toes, though only the inner three (or in some cases four) bore claws. Their forelimbs were rather more slender and ended in pillar-like hands built for supporting weight; only the thumb bore a claw. Many illustrations of sauropods in the flesh miss these facts, inaccurately depicting sauropods with hooves capping the claw-less digits of the feet, and/or multiple claws or hooves on the hands.
Size
The sauropods' most defining characteristic was their size. Even the dwarf sauropods like Europasaurus (perhaps 5 to 6 metres, or 20 feet long) were counted among the largest animals in their ecosystem. Their only real competitors in terms of size are the rorqual whales, such as the Blue Whale. But unlike whales, sauropods were primarily land-based animals.
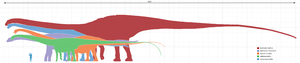
Their body design did not vary as much as other dinosaurs, perhaps due to size constraints, but they still displayed ample variety. Some, like the diplodocids, possessed tremendously long tails which they may have been able to crack like a whip to deter or even injure predators,[2] or to make sonic booms.[3] Supersaurus, at 33 to 34 metres (108 to 112 ft) long,[4] is the longest sauropod known from reasonably complete remains, but others, like the old record holder, Diplodocus, are still extremely long. The holotype (and now lost) vertebra of Amphicoelias fragillimus may have come from an animal 58 metres (190 ft) long;[5] its vertebral column would have been substantially longer than that of the blue whale. The longest terrestrial animal alive today, the reticulated python, only reaches lengths of 8.7 metres (29 ft).[6]
Others, like the brachiosaurids, were extremely tall, with high shoulders and extremely long necks. Sauroposeidon is probably the tallest, reaching about 18 metres (60 ft) high, with the previous record for longest neck being held by Mamenchisaurus. By comparison the giraffe, the tallest of all living animals, is only 4.8 to 5.5 metres (16 to 18 ft) tall.
Some were almost incredibly massive: Argentinosaurus is probably the heaviest at 80 to 100 metric tonnes (90 to 110 tons), though Paralititan, Andesaurus, Antarctosaurus, and Argyrosaurus are of comparable sizes. There is some very poor evidence of an even more massive titanosaurian, Bruhathkayosaurus, which might have weighed between 175 to 220 tonnes (190 to 240 tons). The largest land animal alive today, the Savannah elephant, weighs no more than 10 metric tons (11 short tons).
Among the smallest sauropods were the primitive Ohmdenosaurus (4 m, or 13 ft long), the dwarf titanosaur Magyarosaurus (5.3 m or 17 ft long), and the dwarf brachiosaurid Europasaurus, which was 6.2 meters long as a fully-grown adult.[7] Its small stature was probably the result of insular dwarfism of a herd of sauropods stranded on an island in what is now Germany. Also notable is the diplodocoid sauropod Brachytrachelopan, which was the shortest member of its group thanks to its unusually short neck. Unlike other sauropods, whose necks could grow to up to four times the length of their backs, the neck of Brachytrachelopan was shorter than its backbone.
Limbs and feet
As massive quadrupeds, sauropods developed specialized graviportal (weight-bearing) limbs. The hind feet were broad, and retained three claws in most species[8]. Particularly unusual compared with other animals were the highly modified front feet (manus). The front feet of sauropods were very dissimilar from those of modern large quadrupeds such as elephants. Rather than splaying out to the sides to create a wide foot as in elephants, the manus bones of sauropods were arranged in fully vertical columns, with extremely reduced finger bones (though it is not clear if the most primitive sauropods, such as Vulcanodon and Barapasaurus, had such forefeet)[9]. The front feet were so modified in eusauropods that individual digits would not have been visible in life.
The arrangement of the forefoot bone (metacarpal) columns in eusauropods was semi-circular, so sauropod forefoot prints are horseshoe-shaped. Unlike elephants, print evidence shows that sauropods lacked any fleshy padding to back the front feet, making them concave.[9] The only claw visible in most sauropods was the distinctive thumb claw (associated with digit I). Almost all sauropods had such a claw, though what purpose it served is unknown. The claw was largest (as well as tall and laterally flattened) in diplodocids, and very small in brachiosaurids, some of which seem to have lost the claw entirely based on trackway evidence.[10]
Titanosaurs may have lost the thumb claw completely (with the exception of early forms such as Janenschia). Titanosaurs were most unusual among sauropods, as in addition to the external claw, they completely lost the digits of the front foot. Advanced titanosaurs had no digits or digit bones, and walked only on horseshoe-shaped "stumps" made up of the columnar metacarpal bones.[11]
Print evidence from Portugal shows that in at least some sauropods (probably brachiosaurids), the bottom and sides of the forefoot column was likely covered in small, spiny scales, which left score marks in the prints.[12] In titanosaurs, the ends of the metacarpal bones that contacted the ground were unusually broad and squared-off, and some specimens preserve the remains of soft tissue covering this area, suggesting that the front feet were rimmed with some kind of padding in these species.[11]
Matthew Bonnan[13][14] has shown that sauropod dinosaur long bones grew isometrically: that is, there was little to no change in shape as juvenile sauropods became gigantic adults. Bonnan suggested that this odd scaling pattern (most vertebrates shown significant shape changes in long bones associated with increasing weight support) might be related to a stilt-walker principle (suggested by amateur scientist Jim Schmidt) in which the long legs of adult sauropods allowed them to easily cover great distances without changing their overall mechanics.
Tail
The book version of Walking with Dinosaurs expresses a theory that Diplodocus could crack its tail like a stockwhip to make a loud noise for communication.
Air sacs
Along with other saurischian dinosaurs (such birds and other theropods), sauropods had a system of air sacs, evidenced by indentations and hollow cavities in most of their vertebrae. Pneumatic, hollow bones are a characteristic feature of all sauropods.[15]
The bird-like hollowing of sauropod bones was recognized early in the study of these animals, and in fact at least one sauropod specimen found in the 19th century (Ornithopsis) was originally misidentified as a pterosaur (a flying reptile) because of this.[16]
Armor
Some sauropods had armor. There were genera with spined backs, such as Agustinia, and some had small clubs on their tails, like Shunosaurus. Several titanosaurs, such as Saltasaurus and Ampelosaurus, had small bony osteoderms covering portions of their bodies.
Palaeobiology
Herding and parental care

Many lines of fossil evidence, from both bone beds and trackways, indicate that sauropods were gregarious animals that formed herds. However, the makeup of the herds varied between species. Some bone beds, for example a site from the Middle Jurassic of Argentina, appear to show herds made up of individuals of various age groups, mixing juveniles and adults. However, a number of other fossil sites and trackways indicate that many sauropod species travelled in herds segregated by age, with juveniles forming herds separate from adults. Such segregated herding strategies have been found in species such as Alamosaurus, Bellusaurus, and some diplodocids.[17]
In a review of the evidence for various herd types, Myers and Fiorillo attempted to explain why sauropods appear to have often formed segregated herds. Studies of microscopic tooth wear show that juvenile sauropods had diets that differed from their adult counterparts. Differing tooth wear suggests that juveniles had different feeding strategies than adults, so herding together would not have been as productive as herding separately, where individual herd members could forage in a coordinated way. The vast size difference between juveniles and adults may also have played a part in the different feeding and herding strategies.[17]

Since the segregation of juveniles and adults must have taken place soon after hatching, Myers and Fiorillo concluded that species with age-segregated herds could not have exhibited much parental care, if any.[17] On the other hand, scientists who have studied age-mixed sauropod herds suggested that these species may have cared for their young for an extended period of time before the young reached adulthood.[18]
Exactly how segregated versus age-mixed herding varied across different groups of sauropods is unknown. Further examples of gregarious behavior will need to be discovered from more sauropod species to begin detecting possible patterns of distribution.[17]
Rearing stance

Since early in the history of their study, scientists such as Osborn have speculated that sauropods could rear up on their hind legs, using the tail as the third 'leg' of a tripod.[19] A skeletal mount depicting the diplodocid Barosaurus lentus rearing up on its hind legs at the American Museum of Natural History is one illustration of this hypothesis. In a 2005 paper, Rothschild and Molnar reasoned that if sauropods had adopted a bipedal posture at times, there would be evidence of stress fractures in the forelimb 'hands'. However, none were found after they examined a large number of sauropod skeletons.[20]

Heinrich Mallison (in 2009) was the first to study the physical potential for various sauropods to rear into a tripodal stance. Mallison found that some characters previously linked to rearing adaptations were actually unrelated (such as the wide-set hip bones of titanosaurs) or would actually have hindered rearing. For example, titanosaurs had an unusually flexible backbone, which would have decreased stability in a tripodal posture and would have put more strain on the muscles. Likewise, it is unlikely that brachiosaurids could rear up onto the hind legs, given their center of gravity was much farther forward than other sauropods, which would cause such a stance to be unstable.[21]
Diplodocids, on the other hand, appear to have been well adapted for rearing up into a tripodal stance. Diplodocids had a center of mass directly over the hips, giving them greater balance on two legs. Diplodocids also had the most mobile necks of sauropods, a well-muscled pelvic girdle, and tail vertebrae with a specialised shape that would allow the tail to bear weight at the point it contacted the ground. Mallison concluded that diplodocids were better adapted to rearing than elephants, which do so occasionally in the wild. He also argues that stress fractures in the wild do not occur from everyday behaviour[21], such as feeding-related activities (contra Rothschild and Molnar)[20].
Head and neck posture
There is controversy over whether sauropods held their heads near vertically or horizontally. The claim that the long necks of sauropods were used for browsing high trees has been questioned on the basis of calculations of the energy needed to create the arterial blood pressure for the head if it was held upright. These calculations suggest this would have taken up roughly half of its energy intake.[22] Further, to supply blood to the head vertically held high would have required blood pressure of around 700 mmHg at the heart. This would have required that their hearts were 15 times the size of whales of similar size.[23] This suggests it was more likely the long neck was usually held horizontally to enable them to feed on plants over a very wide area without needing to move their bodies—a potentially large saving in energy for 30 to 40 ton animals. In support of this, reconstructions of the necks of Diplodocus and Apatosaurus show that they are basically straight with a gentle decline orientating their heads in a "neutral, undeflected posture" when close to ground.[24]
However, research on living animals has suggested that sauropod heads were held in an upright S-shaped curve. Inference from bones about "neutral head postures" which suggest a horizontal position[24] may be unreliable, according to this research. If applied to living animals it would imply they also held their heads in this position, even though they in fact do not.[25][26]
Trackways and locomotion

Sauropod trackways and other fossil footprints (known as "ichnites") are known from abundant evidence present on most continents. Ichnites have helped support other biological hypotheses about sauropods, including general fore and hind foot anatomy (see Limbs and feet above). Generally, prints from the forefeet are much smaller than the hind feet, and often crescent-shaped. Occasionally ichnites preserve traces of the claws, and help confirm which sauropod groups lost claws or even digits on their forefeet.[27]
Generally, sauropod trackways are divided into three categories based on the distance between opposite limbs: narrow gauge, medium gauge, and wide gauge. The gauge of the trackway can help determine how wide-set the limbs of various sauropods were and how this may have impacted the way they walked.[27] A 2004 study by Day and colleagues found that a general pattern could be found among groups of advanced sauropods, with each sauropod family being characterised by certain trackway gauges. They found that most sauropods other than titanosaurs had narrow-gauge limbs, with strong impressions of the large thumb claw on the forefeet. Medium gauge trackways with claw impressions on the forefeet probably belong to brachiosaurids and other primitive titanosauriformes, which were evolving wider-set limbs but retained their claws. Primitive true titanosaurs also retained their forefoot claw but had evolved fully wide gauge limbs. Wide gauge limbs were retained by advanced titanosaurs, trackways from which show a wide gauge and lack of any claws or digits on the forefeet.[28]
Size evolution
Several scientists have attempted to address the question of why sauropods attained such huge sizes. Gigantic sizes were reached early in sauropod evolution, going back to the first true sauropods in the late Triassic Period. According to Kenneth Carpenter, whatever evolutionary pressure caused large size must have therefore been present from the early origins of the group.[5]
Studies of mammalian herbivores that attained large size, such as elephants, have found that larger size in plant-eating animals leads to greater efficiency in digesting food. Since larger animals have longer digestive systems, food is kept in digestion for significantly longer periods of time, allowing large animals to survive on lower-quality food sources. This is especially true of animals with a large number of 'fermentation chambers' along the intestine which allow microbes to accumulate and ferment plant material, aiding digestion. Throughout their evolutionary history, sauropod dinosaurs were found primarily in semi-arid, seasonally dry environments, with a corresponding seasonal drop in the quality of food during the dry season. The environment of most gigantic Late Jurassic sauropods such as Amphicoelias was essentially a savanna, similar to the arid environments in which modern giant herbivores are found, supporting the idea that poor-quality food in an arid environment promotes the evolution of giant herbivores. Carpenter argued that other benefits of large size, such as relative immunity from predators, lower energy expenditure, and longer life span, were probably secondary advantages, and that sauropods attained large size primarily to help process food more efficiently.[5]
History of discovery
The first scrappy fossil remains now recognized as sauropods all came from England and were originally interpreted in a variety of different ways. Their relationship to other dinosaurs was not recognized until well after their initial discovery.


The first sauropod fossil to be scientifically described was a single tooth known by the non-Linnaean descriptor Rutellum implicatum.[29] This fossil was described by Edward Lhuyd in 1699, but was not recognized as a giant prehistoric reptile at the time.[30] Dinosaurs would not be recognized as a group until over a century later.
Richard Owen published the first modern scientific description of sauropods in 1841, in his paper naming Cetiosaurus and Cardiodon. Cardiodon was known only from a two unusual, heart-shaped teeth (from which it got its name), which could not be identified beyond the fact that they came from a previously unknown large reptile. Cetiosaurus was known from slightly better, but still scrappy remains. Owen thought at the time that Cetiosaurus was a giant marine reptile related to modern crocodiles, hence its name, which means "whale lizard". A year later, when Owen coined the name Dinosauria, he did not include Cetiosaurus and Cardiodon in that group.[31]
In 1850, Gideon Mantell recognized the dinosaurian nature of several bones assigned to Cetiosaurus by Owen. Mantell noticed that the leg bones contained a medullary cavity, a characteristic of land animals. He assigned these specimens to the new genus Pelorosaurus, and grouped it together with the dinosaurs. However, Mantell still did not recognize the relationship to Cetiosaurus.[16]
The next sauropod find to be described and misidentified as something other than a dinosaur were a set of hip vertebrae described by Harry Seeley in 1870. Seeley found that the vertebrae were very lightly constructed for their size and contained openings for air sacs (pneumatization). Such air sacs were at the time known only in birds and pterosaurs, and Seeley considered the vertebrae to come from a pterosaur. He named the new genus Ornithopsis, or "bird face" because of this.[16]
When more complete specimens of Cetiosaurus were described by Phillips in 1871, he finally recognized the animal as a dinosaur related to Pelorosaurus.[32] However, it was not until the description of new, nearly complete sauropod skeletons from the United States (representing Apatosaurus and Camarasaurus) later that year that a complete picture of sauropods emerged. An approximate reconstruction of a complete sauropod skeleton was produced by John A. Ryder, based on the remains of Camarasaurus, though many features were still inaccurate or incomplete according to later finds and biomechanical studies.[33] Also in 1877, Richard Lydekker named another relative of Cetiosaurus, Titanosaurus, based on an isolated vertebra.[16]
In 1878, the most complete sauropod yet was found and described by Othniel Charles Marsh, who named it Diplodocus. With this find, Marsh also created a new group to contain Diplodocus, Cetiosaurus, and their increasing roster of relatives to differentiate them from the other major groups of dinosaurs. Marsh named this group Sauropoda, or "lizard feet".[16]
Classification
Classification of the sauropods has largely stabilised in recent years, though there are still some uncertainties, such as the placement of Euhelopus, Haplocanthosaurus, Jobaria and Nemegtosauridae. The following are two alternative recent classifications (showing supra-generic clades only in the second example). These are by no means an exhaustive list of recent sauropod classification schemes. In some cases, families like Vulcanodontidae, Cetiosauridae and Omeisauridae are not included because they are considered paraphyletic, or even (in the case of Camarasauridae) polyphyletic.
Taxonomy
This taxonomy follows Wilson & Sereno 1998, Yates 2003 and in press,[34] Galton 2001,[1] and Wilson 2002,[35] with ranks after Benton, 2004.[36]
- Infraorder Sauropoda
- Isanosaurus
- Kotasaurus
- Lessemsaurus
- Family Blikanasauridae
- Family Melanorosauridae
- Family Vulcanodontidae
- Family Cetiosauridae
- Family Omeisauridae
- ?Family Tendaguridae
- Clade Turiasauria
- Division Neosauropoda
- Haplocanthosaurus
- ?Jobaria
- Superfamily Diplodocoidea
- Family Rebbachisauridae
- Family Dicraeosauridae
- Family Diplodocidae
- Subdivision Macronaria
- Family Brachiosauridae
- Family Camarasauridae
- Family Euhelopodidae
- Superfamily Titanosauroidea
Phylogeny
Cladogram simplified after Wilson, 2002.[35]
| Sauropoda |
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
References
- ↑ Marsh, O.C. (1878). "Principal characters of American Jurassic dinosaurs. Part I". American Journal of Science and Arts 16: 411-416
- ↑ Bakker, Robert (1994). "The Bite of the Bronto" Earth 3:(6):26–33.
- ↑ Peterson, Ivars (March 2000). "Whips and Dinosaur Tails". Science News. http://www.sciencenews.org/articles/20000318/mathtrek.asp. Retrieved 2007-07-07.
- ↑ Lovelace, David M.; Hartman, Scott A.; and Wahl, William R. (2007). "Morphology of a specimen of Supersaurus (Dinosauria, Sauropoda) from the Morrison Formation of Wyoming, and a re-evaluation of diplodocid phylogeny". Arquivos do Museu Nacional 65 (4): 527–544.
- ↑ 5.0 5.1 5.2 Carpenter, K. (2006). "Biggest of the big: a critical re-evaluation of the mega-sauropod Amphicoelias fragillimus." In Foster, J.R. and Lucas, S.G., eds., 2006, Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin 36: 131–138.
- ↑ Murphy JC, Henderson RW. 1997. Tales of Giant Snakes: A Historical Natural History of Anacondas and Pythons. Krieger Pub. Co. 221 pp. ISBN 0894649957.
- ↑ Sander, P.M., Mateus, O., Laven, T. & Knötschke, N. (2006). "Bone histology indicates insular dwarfism in a new Late Jurassic sauropod dinosaur". Nature, 441: 739-741.
- ↑ Bonnan, M.F. 2005. Pes anatomy in sauropod dinosaurs: implications for functional morphology, evolution, and phylogeny; pp. 346-380 in K. Carpenter and V. Tidwell (eds.), Thunder-Lizards: The Sauropodomorph Dinosaurs. Bloomington, IN: Indiana University Press.
- ↑ 9.0 9.1 Bonnan, M.F. 2003. The evolution of manus shape in sauropod dinosaurs: implications for functional morphology, forelimb orientation, and sauropod phylogeny. Journal of Vertebrate Paleontology, 23(3): 595-613.
- ↑ Upchurch, P. (1994). "Manus claw function in sauropod dinosaurs". Gaia, 10: 161-171.
- ↑ 11.0 11.1 Apesteguía, S. (2005). "Evolution of the titanosaur metacarpus". Pp. 321-345 in Tidwell, V. and Carpenter, K. (eds.) Thunder-Lizards: The Sauropodomorph Dinosaurs. Indianapolis: Indiana University Press.
- ↑ Milàn, J., Christiansen, P. and Mateus, O. (2005). "A three-dimensionally preserved sauropod manus impression from the Upper Jurassic of Portugal: implications for sauropod manus shape and locomotor mechanics". Kaupia, 14: 47-52.
- ↑ Bonnan, M.F. 2004. Morphometric analysis of humerus and femur shape in Morrison sauropods: implications for functional morphology and paleobiology. Paleobiology, 30(3): 444-470.
- ↑ Bonnan, M.F. 2007. Linear and geometric morphometric analysis of long bone scaling patterns in Jurassic Neosauropod dinosaurs: their functional and paleobiological implications. The Anatomical Record, 290(9): 1089-1111.
- ↑ Wedel, M.J. (2009). "Evidence for bird-like air sacs in Saurischian dinosaurs". (pdf) Journal of Experimental Zoology, 311A: 18pp.
- ↑ 16.0 16.1 16.2 16.3 16.4 Taylor, M.P. (In press). "Sauropod dinosaur research: a historical review". In Richard Moody, Eric Buffetaut, David M. Martill and Darren Naish (eds.), Dinosaurs (and other extinct saurians): a historical perspective. HTML abstract.
- ↑ 17.0 17.1 17.2 17.3 Myers, T.S. and Fiorillo, A.R. (2009). "Evidence for gregarious behavior and age segregation in sauropod dinosaurs". Palaeogeography, Palaeoclimatology, Palaeoecology, 274: 96-104.
- ↑ Coria, R.A. (1994). "On a monospecific assemblage of sauropod dinosaurs from Patagonia: implications for gregarious behavior". GAIA, 10: 209–213.
- ↑ Osborn, H.F. (1899). "A skeleton of Diplodocus, recently mounted in the American Museum". Science, new series, vol. 10: 870-874.
- ↑ 20.0 20.1 Rothschild, B.M. and Molnar, R.E. (2005). "Sauropod Stress Fractures as Clues to Activity". In Carpenter, K. and Tidswell, V. (eds.). Thunder Lizards: The Sauropodomorph Dinosaurs. Indiana University Press. pp. 381–391. ISBN 0-253-34542-1.
- ↑ 21.0 21.1 Mallison, H. (2009). "Rearing for food? Kinetic/dynamic modeling of bipedal/tripodal poses in sauropod dinosaurs". P. 63 in Godefroit, P. and Lambert, O. (eds), Tribute to Charles Darwin and Bernissart Iguanodons: New Perspectives on Vertebrate Evolution and Early Cretaceous Ecosystems. Brussels.
- ↑ Seymour, R.S. (2009). "Raising the sauropod neck: it costs more to get less". Biol Lett. 23;5(3):317-9. PMID 19364714
- ↑ Seymour, R.S. and Lillywhite, H.B. (2000). "Hearts, neck posture and metabolic intensity of sauropod dinosaurs". Proc Biol Sci. 22;267(1455):1883-7. PMID 11052540
- ↑ 24.0 24.1 Stevens, K.A. and Parrish, J.M. (1999). "Neck posture and feeding habits of two Jurassic sauropod dinosaurs". Science. 284(5415):798-800. PMID 10221910
- ↑ Taylor, M.P., Wedel, M.J., and Naish, D. (2009). "Head and neck posture in sauropod dinosaurs inferred from extant animals". Acta Palaeontologica Polonica 54 (2), 2009: 213-220 abstract
- ↑ Museums and TV have dinosaurs' posture all wrong, claim scientists. Guardian, 27 May 2009
- ↑ 27.0 27.1 Riga, B.J.G. and Calvo, J.O. (2009). "A new wide-gauge sauropod track site from the Late Cretaceous of Mendoza, Neuquen Basin, Argentina". Paleontology, 52(3): 631-640.
- ↑ Day, J.J., Norman, D.B., Gale, A.S., Upchurch, P. and Powell, H.P. (2004). "A Middle Jurassic dinosaur trackway site from Oxfordshire, UK". Palaeontology, 47: 319–348.
- ↑ Delair, J.B., and Sarjeant, W.A.S. (2002). "The earliest discoveries of dinosaurs: the records re-examined". Proceedings of the Geologists' Association, 113: 185-197.
- ↑ Lhuyd, E. (1699). Lithophylacii Britannici Ichnographia, sive lapidium aliorumque fossilium Britannicorum singulari figura insignium. Gleditsch and Weidmann: London.
- ↑ Owen, R. (1842). "Report on British Fossil Reptiles". Part II. Report of the British Association for the Advancement of Science, Plymouth, England.
- ↑ Phillips, J. (1871). Geology of Oxford and the Valley of the Thames. Oxford: Clarendon Press, 523 pp.
- ↑ Osborn, H.F., and Mook, C.C. (1921). "Camarasaurus, Amphicoelias and other sauropods of Cope". Memoirs of the American Museum of Natural History, n.s. 3:247-387 and plates LX-LXXXV.
- ↑ Yates, Adam M. (in press). "A revision of the problematic sauropodomorph dinosaurs from Manchester, Connecticut and the status of Anchisaurus Marsh". Palaeontology in press: no. doi:10.1111/j.1475-4983.2010.00952.x.
- ↑ 35.0 35.1 Wilson, J. A. (2002). "Sauropod dinosaur phylogeny: critique and cladistic analysis". Zoological Journal of the Linnean Society, 136: 217-276.
- ↑ Benton, M.J. (2004). Vertebrate Palaeontology, Third Edition. Blackwell Publishing, 472 pp.
- Bob Strauss, 2008, Sauropods: The Biggest Dinosaurs that Ever Lived, The New York Times
- Kristina Curry Rogers and Jeffrey A. Wilson, 2005, The Sauropods: Evolution and Paleobiology, University of California Press, Berkeley, ISBN 0-520-24623-3
- Upchurch, P., Barrett, P.M. and Dodson, P. 2004. Sauropoda. In The Dinosauria, 2nd edition. D. Weishampel, P. Dodson, and H. Osmólska (eds.). University of California Press, Berkeley. Pp. 259–322.