Spermatozoon
| Spermat | |
|---|---|
 |
|
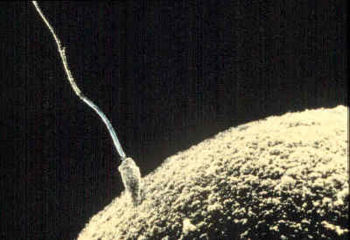
| A sperm cell attempts to penetrate an ovum coat to fertilize it. | |
 |
|
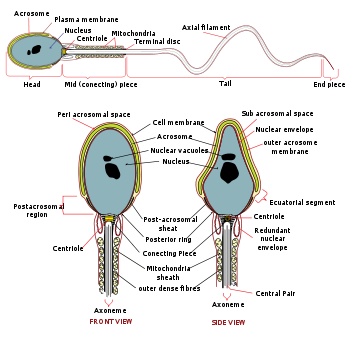
| Diagram of a human spermatozoon | |
| MeSH | Spermatozoa |
| Dorlands/Elsevier | Spermatozoon |
A sperm, from the ancient Greek word σπέρμα (seed) and ζῷον (living being) and more commonly known as a sperm cell, is the haploid cell that is the male gamete. It joins an ovum to form a zygote. A zygote is a single cell, with a complete set of chromosomes, that normally develops into an embryo.
Sperm cells contribute half of the genetic information to the diploid offspring. In mammals, the sex of the offspring is determined by the sperm cell: a spermatozoon bearing a Y chromosome will lead to a male (XY) offspring, while one bearing an X chromosome will lead to a female (XX) offspring (the ovum always provides an X chromosome). Sperm cells were first observed by Anton van Leeuwenhoek in 1677.[1]
Contents |
Mammalian spermatozoan structure, function, and size
Humans
The human sperm cell is the reproductive cell in males. Sperm cells come in two types; "male" and "female". Sperm cells that give rise to female (XX) offspring after fertilization differ in that they carry an X chromosome, while sperm cells that give rise to male (XY) offspring carry a Y chromosome. The "female" sperm cell also differs phenotypically in that it has a larger head in comparison to the "male" sperm cell .
In male humans, sperm cells consists of a head 5 µm by 3 µm and a tail 50 µm long. The Reynolds number associated with spermatozoa is in the order of 1×10-2, so it is known that the spermatozoa exhibit non-turbulent flow. The tail flagellates, which propels the sperm cell (at about 1–3 mm/minute in humans) by whipping in an elliptical cone.[2]. Semen has an alkaline nature, and they do not reach full motility (hypermotility) until they reach the vagina where the alkaline pH is neutralized by acidic vaginal fluids. This gradual process takes 20–30 minutes. In this time, fibrinogen from the seminal vesicles forms a clot, securing and protecting the sperm. Just as they become hypermotile, fibrinolysin from the prostate dissolves the clot, allowing the sperm to progress optimally.
The spermatozoon is characterized by a minimum of cytoplasm and the most densely packed DNA known in eukaryotes. Compared to mitotic chromosomes in somatic cells, sperm DNA is at least sixfold more highly condensed.[3]
During fertilization, the sperm's mitochondria are destroyed by the egg cell, and this means only the mother is able to provide the baby's mitochondria and mitochondrial DNA, which have an important application in tracing maternal ancestry. However it has been recently discovered that mitochondrial DNA can be recombinant [4].
Avoidance of immune system response
Glycoprotein molecules on the surface of sperm cells are recognised by all human immune systems, and interpreted as a signal that the cell should not be rejected. The male immune system might otherwise attack sperm whilst in the testes, and the female immune system would attack sperm in the reproductive tract. The specific glycoproteins coating sperm cells are also utilized by some cancerous and bacterial cells, some parasitic worms, and HIV-infected white blood cells, in order to avoid an immune response from the host organism.[5]
The blood-testis barrier, maintained by the tight junctions between the Sertoli cells of the seminiferous tubules, prevents communication between the forming spermatozoa and the blood vessels within the interstitial space. This gives the forming spermatozoa an immunological advantage and thus prevents them from eliciting an immune response. The blood-testis barrier is also important in preventing toxic substances from disrupting spermatogenesis.
Spermatozoa in other organisms

- See also: Sperm
Animals
Fertilization relies on spermatozoa for most sexually reproductive animals.
The fruit fly[3] has the largest known spermatozoon relative to its size. Drosophila melanogaster produces sperm that can be up to 1.8 mm in size, which is longer than the adult fly. The incredibly long tail is thought to block other sperm from entering the egg. The entire sperm, tail included, gets incorporated into the oocyte cytoplasm.[6]
The wood mouse Apodemus slvaticus possesses spermatozoa with falciform morphology. What makes these gametocytes even more unique is the presence of an apical hook on the sperm head. This hook is used to attach to the hooks or to the flagella of other spermatozoa. Aggregation is caused by these attachments and mobile trains result. These trains provide improved motility in the female reproductive tract and are a means by which fertilization is promoted. [7]
Sea urchins such as Arbacia punctulata—are the workhorses of sperm research, because they spawn large numbers of sperm into the sea, making them well-suited as model organisms for research experiments.
Plants, algae and fungi
The gametophytes of bryophytes, ferns and some gymnosperms produce motile sperm cells, contrary to pollen grains employed in most gymnosperms and all angiosperms. This renders sexual reproduction in the absence of water impossible, since water is a necessary medium for sperm and egg to meet. Algae and lower plant sperm cells are often multi-flagellated (see image) and thus morphologically different from animal spermatozoa.
Some algae and fungi produce non-motile sperm cells, called spermatia. In higher plants and some algae and fungi, fertilization involves the migration of the sperm nucleus through a fertilization tube (e.g. pollen tube in higher plants) to reach the egg cell.
Spermatozoa production in mammals
Spermatozoa are produced in the seminiferous tubules of the testes in a process called spermatogenesis. Round cells called spermatogonia divide and differentiate eventually to become spermatozoa. During copulation the cloaca or vagina gets inseminated, and then the spermatozoa move through chemotaxis to the ovum inside a Fallopian tube or the uterus.
Spermatozoa Activation

Mammalian sperm cells become even more active when they approach an egg cell in a process called sperm activation. Sperm activation has been shown to be caused by calcium ionophores in vitro, progesterone released by nearby cumulus cells and binding to ZP3 of the zona pellucida.
The initial change is called "hyperactivation", which causes a change in spermatozoa motility. They swim faster and their tail movements become more forceful and erratic.
A recent discovery links hyperactivation to a sudden influx of calcium ion into the tails. The whip-like tail (flagellum) of the sperm is studded with ion channels formed by proteins called CatSper. These channels are selective, allowing only calcium ion to pass. The opening of CatSper channels is responsible for the influx of calcium. The sudden rise in calcium levels causes the flagellum to form deeper bends, propelling the sperm more forcefully through the viscous environment. Sperm hyperactivity is necessary for breaking through two physical barriers that protect the egg from fertilization.
The second process in sperm activation is the acrosome reaction. This involves releasing hyaluronidase to digest cumulus cells surrounding the oocyte and exposing acrosin attached to the inner membrane of the sperm. The cumulus cells are embedded in a gel-like substance made primarily of hyaluronic acid, and developed in the ovary with the egg and support it as it grows.
Acrosin digests the zona pellucida and membrane of the oocyte. Part of the sperm's cell membrane then fuses with the egg cell's membrane, and the contents of the head sink into the egg. ZP3, one of the proteins that make up the zona pellucida, binds to a partner molecule on the sperm. This lock-and-key type mechanism is species-specific and prevents the sperm and egg of different species from fusing. There is some evidence that this binding is what triggers the acrosome to release the enzymes that allow the sperm to fuse with the egg.
Upon penetration, the oocyte is said to have become activated. It undergoes its secondary meiotic division, and the two haploid nuclei (paternal and maternal) fuse to form a zygote. In order to prevent polyspermy and minimise the possibility of producing a triploid zygote, several changes to the egg's cell membranes renders them impenetrable shortly after the first sperm enters the egg.
References
- ↑ "Timeline: Assisted reproduction and birth control". Retrieved on 2006-04-06.
- ↑ Sumio Ishijima, Shigeru Oshio, Hideo Mohri, "Flagellar movement of human spermatozoa", Gamete research, 1986, vol. 13, no3, pp. 185–197 (27 ref.) [1]
- ↑ Ward WS, Coffey DS (1991). "DNA packaging and organization in mammalian spermatozoa: comparison with somatic cells". Biol. Reprod. 44 (4): 569–74. doi:. PMID 2043729.
- ↑ Marilena D'Aurelio et al., "Heterologous mitochondrial DNA recombination in human cells", Human Molecular Genetics 2004 13(24):3171–3179; doi:10.1093/hmg/ddh326 [2]
- ↑ "Sperm clue to 'disease immunity'", BBC News (2007-12-17).
- ↑ Gilbert, Scott F., Developmental Biology, Eighth Edition. 2006. Sinauer Associates, pp. 254
- ↑ Moore, Harry et al, Exceptional sperm cooperation in Wood Mouse.Nature 418, 174-177 (2002).
External links
- The Handbook of Andrology
- Sperm hyperactivity
- Slower conception 'leads to boys'
- Photos of sperm under a microscope
|
|||||||||||||||||||||||||