Plasmodium falciparum
| Plasmodium falciparum | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
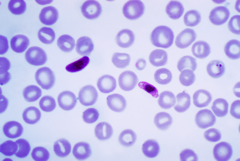 Blood smear of Plasmodium falciparum
|
||||||||||||||
| Scientific classification | ||||||||||||||
|
||||||||||||||
| Binomial name | ||||||||||||||
| Plasmodium falciparum Welch, 1897 |
Plasmodium falciparum is a protozoan parasite, one of the species of Plasmodium that cause malaria in humans. It is transmitted by Anopheles mosquitoes. P. falciparum is the most dangerous of these infections as P. falciparum malaria has the highest rates of complications and mortality. In addition it accounts for 80% of all human malarial infections and 90% of the deaths. It is more prevalent in sub-Saharan Africa than in other regions of the world.
Contents |
Background
Malaria is caused by an infection with protozoa of the genus Plasmodium. The name malaria, from the Italian mala aria, meaning bad air, comes from the linkage suggested by Giovanni Maria Lancisi (1717) of malaria with the poisonous vapours of swamps. This species name comes from the Greek falx meaning sickle-shaped, and parum meaning birth or multiple births. The organism itself was first seen by Laveran on November 6th 1880 at a military hospital in Constantine, Algeria, when he discovered a microgametocyte exflagellating. Manson (1894) hypothesised that mosquitoes could transmit malaria - an association made considerably earlier in India, possibly as early as 2000BC. This hypothesis was experimentally confirmed independently by Giovanni Battista Grassi and Ronald Ross in 1898. Grassi (1900) proposed an exerythrocytic stage in the life cycle and this was later confirmed by Short, Garnham, Covell and Shute (1948) who found Plasmodium vivax in the human liver.
Globally, malaria is the most important parasitic disease of humans and claims the lives of more children worldwide than any other infectious disease. Since 1900 the area of the world exposed to malaria has been halved but in this time two billion more are presently exposed. Morbidity as well as mortality is substantial. Infection rates in children in endemic areas are of the order of 50%: chronic infection has been shown to reduce school scores by up to 15%. Reduction in the incidence of malaria coincides with increased economic output.
While there are no effective vaccines for any of the six or more species that cause human malaria, drugs have been employed for centuries. In 1640, Huan del Vego first employed the tincture of the cinchona bark for treating malaria: the native Indians of Peru and Ecuador had been using it even earlier for treating fevers. Thompson (1650) introduced this "Jesuits' bark" to England: its first recorded use there was by Dr John Metford of Northampton in 1656. Morton (1696) presented the first detailed description of the clinical picture of malaria and of its treatment with cinchona. Gize (1816) studied the extraction of crystalline quinine from the cinchona bark and Pelletier and Caventou (1820) in France extracted pure quinine alkaloids which they named quinine and cinchonine.
Plasmodium Life Cycle
- See also: Plasmodium falciparum biology
When an infected mosquito bites a human, sporozoites enter the human circulation. These go to and penetrate the liver cells, where they asexually reproduce, via the process of schizogony. This intracellular, asexually dividing form of the parasite is known as a microgamete, which fertilizes the female gametocyte forming a zygote. The zygote develops into an ookinete, which then sticks to the gut wall of the mosquito, moves to the outermost layer of the stomach to form an oocyst.
Treatment and drug resistance
Attempts to make synthetic antimalarials began in 1891. Atabrine was developed in 1928, was used widely throughout the Pacific in World War II but was deeply unpopular because of the yellowing of the skin it caused. In the late 1930s, the Germans developed chloroquine, which went into use in the North African campaigns. Mao Zedong encouraged Chinese scientists to find new antimalarials after seeing the casualties in the Vietnam War. Artemisinin was discovered in the 1970s based on a medicine described in China in the year 340. This new drug became known to Western scientists in the late 1980s and early 1990s and is now a standard treatment. In 1976 P. falciparum was successfully cultured in vitro for the first time which facilitated the development of new drugs substantially.[1]
Vaccination
Although an antimalarial vaccine is urgently needed, infected individuals never develop a sterilizing (complete) immunity, making the prospects for such a vaccine dim. The parasites live inside cells, where they are largely hidden from the immune response. Infection has a profound effect on the immune system including immune suppression. Dendritic cells suffer a maturation defect following interaction with infected erythrocytes and become unable to induce protective liver-stage immunity. Infected erythrocytes directly adhere to and activate peripheral blood B cells from nonimmune donors. The var gene products, a group of highly expressed surface antigens, bind the Fab and Fc fragments of human immunoglobulins in a fashion similar to protein A to Staphylococcus aureus and this may offer some protection to the parasite from the human immune system. Despite the poor prospects for a fully protective vaccine, it may be possible to develop a vaccine that would reduce the severity of malaria for children living in endemic areas.
Microscopic appearance

Among medical professionals, the preferred method to diagnose malaria and determine which species of Plasmodium is causing the infection is by examination of a blood film microscopically in a laboratory. Each species has distinctive physical characteristics that are apparent under a microscope. In P. falciparum, only early trophozoites and gametocytes are seen in the peripheral blood. It is unusual to see mature trophozoites or schizonts in peripheral blood smears as these are usually sequestered in the tissues. The parasitised erythrocytes are not enlarged and it is common to see cells with more than one parasite within them (multiply parasitised erythrocytes). Occasionally, faint comma-shaped red dots are seen on the red cell surface called "Maurer's dots". The comma shaped dots can also appear as pear shaped blotches.
Plasmodium and the human genome
In the 50,000 years since Plasmodium first infected humans , the presence of the parasite in human populations has altered the human genome in a multitude of ways, as humans have been forced to develop resistance to the disease. Beet, a doctor working in Southern Rhodesia (now Zimbabwe) in 1948, first suggested that sickle-cell disease could offer some protection to malaria. This suggestion was reiterated by J. B. S. Haldane in 1949 who suggested that thalassaemia could provide similar protection. This hypothesis has since been confirmed and has been extended to hemoglobin C and hemoglobin E, abnormalities in ankyrin and spectrin (ovalocytosis , elliptocytosis), in glucose-6-phosphate dehydrogenase deficiency and pyruvate kinase deficiency, loss of the Gerbich antigen (glycophorin C) and the Duffy antigen on the erythrocytes, thalassemias and variations in the major histocompatibility complex classes 1 and 2 and CD32 and CD36.
In 1995 a consortium - the malaria genome project (MGP) - was set up to sequence the genome of P. falciparum. The genome of the parasite mitochondrion was reported in 1995, that of the plastid (apicoplast) in 1996, and the sequence of the first nuclear chromosome (Chromosome 2) in 1998. The sequence of chromosome 3 was reported in 1999 and the entire genome on 3rd October 2002. The ~24 megabase genome is extremely AT rich (~80%) and is organised into 14 chromosomes: just over 5300 genes were described.
Evolution of Plasmodium falciparum
The closest relative of Plasmodium falciparum is Plasmodium reichenowi, a parasite of chimpanzees. Molecular clock analyses suggest that P. falciparum is as old as the human line; the two species diverged at the same time as humans and chimpanzees[2]. However, low levels of polymorphism within the P. falciparum genome suggest a much more recent origin[3]. It may be that this discrepancy exists because P. falciparum is old, but its population recently underwent a great expansion[4].
P. falciparum and P. reichenowi are not closely related to the other Plasmodium species that parasitize humans, or indeed mammals generally. It has been argued that these two species originated from a parasite of birds[5]. More recent analyses do not support this, however, instead suggesting that the ability to parasitize mammals evolved only once within the genus Plasmodium[6].
More recently, P. falciparum has evolved in response to human interventions. Most strains of malaria can be treated with chloroquine, but P. falciparum has developed resistance to this treatment. A combination of quinine and tetracycline has also been used, but there are strains of P. falciparum that have grown resistant to this treatment as well. Different strains of P. falciparum have grown resistant to different treatments. Often the resistance of the strain depends on where it was contracted. Many cases of malaria that come from parts of the Caribbean and west of the Panama Canal as well as the Middle East and Egypt can often be treated with chloroquine, since they have not yet developed resistance. Nearly all cases contracted in Africa, India, and southeast Asia have grown resistant to this medication and there have been cases in Thailand and Cambodia in which the strain has been resistant to nearly all treatments. Often the strain grows resistant to the treatment in areas where the use is not as tightly regulated.
Like most Apicomplexa, malaria parasites harbor a plastid similar to plant chloroplasts, which they probably acquired by engulfing (or being invaded by) a eukaryotic alga, and retaining the algal plastid as a distinctive organelle encased within four membranes (see endosymbiotic theory). The apicomplexan plastid, or apicoplast, is an essential organelle, thought to be involved in the synthesis of lipids and several other compounds, and provides an attractive target for antimalarial drug development, particularly in light of the emergence of parasites resistant to chloroquine and other existing antimalarial agents.
Plasmodium falciparum and Sickle Cell Anemia
Individuals with sickle cell anemia and individuals with sickle cell trait do have reduced parasitemia when compared to wild typed individuals for the hemoglobin protein in red blood cells. Studies have shown these genetic deviations of hemoglobin from normal states provide protection against the deadly parasite that causes malaria (Allison, 1954). Of the four malarial parasites, Plasmodium falciparum causes the most fatal and medically severe form. Malaria is prevalent in tropical countries with an incidence of 300 million per year and a mortality rate of 1to 2 million per year. Roughly 50% of all malarial infections are caused by Plasmodium falciparum (Roberts and Janovy Jr., 2005) Upon infection via a bite from an infected Anopheles mosquito, sporozoites devastate the human body by first, infecting the liver. While in the liver, sporozoites undergo asexual development and merozoites are released into the blood stream. The trophozoites further develop and reproduce by invading red blood cells. During the reproduction cycle, Plasmodium falciparum produces up to 40,000 merozoites in one day. Other blood sporozoans, such as Plasmodium vivax, Plasmodium ovale, and Plasmodium malariae, that infect humans and cause malaria do not have such a productive cycle for invasion. The process of bursting red blood cells does not have any symptoms, however destruction of the cells does cause anemia since the bone marrow cannot compensate for the damage. When red blood cells rupture, hemozoin wastes cause cytokine release, chills, and then fever (Roberts and Janovy Jr., 2005).
Plasmodium falciparum trophozoites develop sticky knobs in red blood cell that adheres to endothelial cells in blood vessels, thus evading clearance in the spleen. The acquired adhesive nature of the red blood cells may cause cerebral malaria when sequestered cells prevent oxygenation of the brain. Symptoms of cerebral malaria include impaired consciousness, convulsions, neurological disorder, and coma (Brown University). Additional complications from Plasmodium falciparum induced malaria include advanced immunosupression (Roberts and Janovy Jr., 2005).
Individuals with sickle cell trait and sickle cell anemia are privileged because they have altered sticky knobs. Research by Cholera, R., Brittain, N., Gillrie, M. et al. (2007) has shown that parasitemia (the ability of a parasite to infect) because merozoites of each parasite species that cause malaria invade the red blood cell in three stages: contact, attachment, and endocytosis. Individuals suffering from sickle cell anemia have deformed red blood cells that interfere with the attachment phase and Plasmodium falciparum and the other forms of malaria have trouble with endocytosis.
These individuals have reduced attachment when compared to red blood cells with the normally functioning hemoglobin because of differing protein interactions. Normally, merozoites enter red blood cells through two PfEMP-1 protein dependent interactions. These interactions promote the malaria inflammatory response associated with symptoms of chills and fever. When these proteins are impaired as in sickle cell cases, parasites cannot undergo cytoadherance interactions and cannot infect the cells, therefore sickle cell anemic individuals and individuals carrying the sickle cell trait have lower parasite loads and shorter time for symptoms than individuals expressing normal red blood cells (Mockenhaupt, 2004).
Individuals with sickle cell anemia may also experience greatly reduced symptoms of malaria because Plasmodium falciparum trophozoites cannot bind to hemoglobin in order to form sticky knobs. Without knob binding complexes, which is an exclusive feature of Plasmodium falciparum, red blood cells do not stick to endothelial walls of blood vessels and infected individuals do not experience symptoms such as cerebral malaria (Cholera, R., Brittain, N., Gillrie, M. et al. 2007).
Many may wonder why natural selection has not phased out sickle cell anemia. The answer to that question lies within answers generated by Cholera, R., Brittain, N., Gillrie, M. et al. (2007). Individuals with sickle cell trait are greatly desired in areas where malarial infections are endemic. Malaria kills between 1 and 2 million people per year. It is the leading cause of death amongst children in tropical regions. Individuals with sickle cell deformities are able to fight Plasmodium parasite infections and do not become victims of malarial demise. Therefore, individuals expressing the genes and individuals carrying genes are selected to remain within the population (Allison 1964). To no surprise the incidence of sickle cell anemia match endemic regions for malarial infections.
References
- ↑ Trager, W; J B Jensen (1976-08-20). "Human malaria parasites in continuous culture". Science (New York, N.Y.) 193 (4254): 673–5. doi:. ISSN 00368075. PMID 781840.
- ↑ Escalante, AA; E Barrio, FJ Ayala (1995). "Evolutionary origin of human and primate malarias: evidence from the circumsporozoite protein gene". Molecular Biology and Evolution 12 (4): 616–26. ISSN 07374038. PMID 7659017.
- ↑ Hartl, DH (2004-01). "The origin of malaria: mixed messages from genetic diversity". Nature Reviews: Microbiology 2 (1): 15–22. doi:. ISSN 17401526. PMID 15035005.
- ↑ Joy, DA; Feng X, Mu J, Furuya T, Chotivanich K, Krettli AU, Ho M, Wang A, White NJ, Suh E, Beerli P, Su XZ. (2003-04-11). "Early origin and recent expansion of Plasmodium falciparum". Science 300 (5617): 318–21. doi:. ISSN 01934511. PMID 15035005.
- ↑ Rathore, D; Wahl AM, Sullivan M, McCutchan TF (2001-04-25). "A phylogenetic comparison of gene trees constructed from plastid, mitochondrial and genomic DNA of Plasmodium species". Molecular and Biochemical Parasitology 114 (1): 89–94. doi:. ISSN 01666851. PMID 11356517.
- ↑ Yotoko, KSC; C Elisei (2006-11). "Malaria parasites (Apicomplexa, Haematozoea) and their relationships with their hosts: is there an evolutionary cost for the specialization?". Journal of Zoological Systematics and Evolutionary Research 44 (4): 265–73. doi:. ISSN 09475745.
Sources and further reading
Overview
{kind=link}
Spatial distribution
Blood slides
- Blood forms
- Blood forms
- Blood forms
- Blood forms
- Multiple blood forms
- Female gametocyte
- Female gametocyte
- ncbi.nlm.nih.gov
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Case histories
Pathology due to Plasmodium falciparum
- Brain
- Gross pathology
- Low power H & E stain
- High power H & E stain showing parasite adherence to the vessel walls
{kind=link}
{kind=link}
- Spleen
- Liver
{kind=link}
- Kidney
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Plasmodium falciparum genome data
- Gardner, M.J.; Hall, N.; Fung, E.; White, O.; Berriman, M.; Hyman, R.W.; Carlton, J.M.; Pain, A.; Nelson, K.E.; Bowman, S.; Others, (2002). "Genome sequence of the human malaria parasite Plasmodium falciparum". Nature 419: 498–511. doi:.
- PlasmoDB: The Plasmodium Genome Resource
- GeneDB Plasmodium falciparum
- Genome
- UCSC Plasmodium Falciparum Browser
Other
- Colombian scientists develop computacional tool to detect the plasmodium falciparum (in spanish)
- Allison AC (1954) Protection afforded by sickle-cell trait against subtertian malareal infection. Br Med J 4857:290 –294.
- Allison AC (1964) Polymorphism and Natural Selection in Human Populations Cold Spring Harb Symp Quant Biol 29:137–149.
- Cholera, R, Brittain, N, Gillrie, M, Lopera-Mesa, T, Diakite, S, & Arie, T (2007). Impaired cytoadherence of Plasmodium falciparum-infected erythrocytes containing sickle hemoglobin.
- Mockenhaupt FP, et al. (2004) Limited influence of haemoglobin variants on Plasmodium falciparum msp1 and msp2 alleles in symptomatic malaria. Trans R Soc Trop Med Hyg 98:302–310.
- Roberts, L, & Jonovy, Jr., J (2005). Foundations of Parasitology. New York: McGraw-Hill.