Hesperornithes
| Hesperornithes Fossil range: Late Cretaceous (Albian - Maastrichtian) |
||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
 Hesperornis
|
||||||||||
| Scientific classification | ||||||||||
|
||||||||||
| Families | ||||||||||
|
Enaliornithidae |
||||||||||
| Synonyms | ||||||||||
|
Odontornithes Marsh, 1873 (partim) |
Hesperornithes is an extinct and highly specialized clade of Cretaceous toothed birds. Hesperornithine birds, apparently limited to former aquatic habitats in the Northern Hemisphere, include genera such as Hesperornis, Parahesperornis, Baptornis, Enaliornis, and probably Potamornis, all strong-swimming predatory waterbirds. Many, if not all species were completely flightless. The largest known hesperornithine, described in 1999 and named Canadaga arctica, may have reached a maximum adult length of over 1.5 meters (five feet).
Hesperornithine birds were, strictly speaking, the only Mesozoic dinosaurs to colonize the oceans (technically speaking, all birds are dinosaurs); the aquatic reptiles of their time, such as the ichthyosaurs and plesiosaurs, were not dinosaurs. Hesperornithine birds went extinct at the K-T boundary, along with enantiornithine protobirds, all non-avian dinosaurs, and many other life forms.
Contents |
Anatomy and ecology
Most of what is known about this group rests on analyses of single species, as few provide sufficiently diverse fossils for analysis. In most cases, what holds true for one hesperornithine also applies to the others, as these birds were quite stereotypical and extreme in their autapomorphies.
Although some of the smaller species might have been able to fly, Hesperornis and Baptornis had only vestigial wings. Like living foot-propelled diving birds, the femur and metatarsus were short, whereas the tibia was long. The legs were also set far back on the body, as in loons, grebes or penguins. Hesperornithids must have been powerful swimmers and divers but extremely ungainly on the land, and probably spent little time ashore except to nest (indeed, it has been suggested that they did not incubate their eggs).
Some researchers think that on land they had to slide on their bellies and push with their legs; the hip and knee joints were shaped such that these birds could not move them dorsoventrally, and in a resting position the feet projected sideways from the body, which would have prevented them from walking upright. Whereas Hesperornis had probably lobes of skin on its toes similar to the grebes of today, the toes of Baptornis could not rotate (necessary in lobed feet to reduce drag) and thus were more likely webbed, as in loons. The dense bones of these birds decreased their buoyancy, making diving easier[1].
The beak was long, probably hooked at the tip, and bore a series of simple, sharp teeth which were set into a longitudinal groove. These and probably helped to seize fish, as does the serrated beak of mergansers today; unlike the reptilian teeth of other toothed birds, those of the hesperornithids were unique[2]. Given that teeth are induced by a rather simple genetic mechanism[3], it is even possible that these birds re-evolved them and that some ancestors of the hesperornithids were toothless.
Systematics and evolution
These birds were originally combined with Ichthyornis in the paraphyletic "Odontornithes" by Othniel Charles Marsh, in 1873. In 1875, they were separated as Odontolcae. The group was often considered to be allied to loons and grebes[4], or to the Paleognathae[5]. These similarities, however, as the more recently determined fact that the osteons of their bones - at least in Hesperornis - were arranged in a pattern similar to that in Neognathae[6], are today considered to be due to convergent evolution[7].
Currently, the hesperornithine birds are recognized as a very specialized lineage that is not ancestral or otherwise closely related to the birds of today. Still, their relationship is close enough that they probably diverged from the ancestors of today's birds as late as the earliest Cretaceous.
The discovery that Gansus yumenensis, which lived some time before the main radiation of hesperornithids, was a primitive member of the Ornithurae - that is, birds which had developed the type of pygostyle that is also found in modern birds - suggests that the ancestors of Hesperornithes - which also ornithuran - may have been birds generally similar to Gansus and living some 130-150 mya. Given the tendency of birds that have become flightless to increase in size markedly in just a few million years, the probability of Hesperornithes deriving from an animal at least superficially similar and not too far related to Gansus is quite high. The new fossil material of that species should assist in determining the number of synapomorphies of hesperornithines and other ornithurans, which have until now been muddied by the numerous autapomorphies of the former.
Loss and/or fusion of caudal vertebrae in pygostyle-like structures was a general trend in Cretaceous avian evolution, and a full pygostyle and associated structures may have evolved more than once to similar shapes. The fact that Gansus had non-pneumatized, dense bones, like those of hesperornithines, although it was not a specialized diver, is interesting to note. Similarly, the bone structure of Hesperornis indicates that as opposed to Enantiornithes and in line with other Ornithurae it showed rapid, uninterrupted growth to adult size[1].
The earliest known hesperornithine is the Early Cretaceous Enaliornis although these birds are somewhat tentatively assigned to this group because of the bad preservation of their remains; the majority of hesperornithine taxa are known from the Late Cretaceous. Small hesperornithine bones are known from the freshwater deposits of the Late Cretaceous of the Judith River Group as well as the Hell Creek and Lance Formations as well as Eurasian sites. These birds were about the size of a cormorant or a loon. Less well-verified is the presence of the lineage around Antarctica, which was a temperate, ice-free region in their time.
Systematics
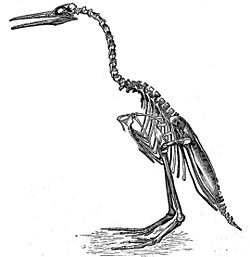
Subclass HESPERORNITHES[8]
This is usually treated as equivalent with Hesperornithiformes. However, given the uncertainties of the evolution of teeth and flightlessness in the Hesperornithes, as noted above, it is here considered a distinct group, so that ancestral taxa do not have to be placed together with more advanced ones.
- Basal and unresolved taxa
- Genus Asiahesperornis (Late Cretaceous) - hesperornithid?
- Genus Judinornis (Late Cretaceous)
- Genus Pasquiaornis (Late Cretaceous)
- Hesperornithes gen. et sp. indet. (Zhuravlovskaya/Zhuravlevo Late Cretaceous of Kushmurun, Kazakhstan)[9]
- Hesperornithes gen. et sp. indet. TMP 89.81.12 (Dinosaur Park Late Cretaceous of Iddesleigh, Canada)[10]
- Order HESPERORNITHIFORMES
- Family Enaliornithidae
- Genus Enaliornis (Early Cretaceous)
- Family Baptornithidae
- Genus Baptornis (Late Cretaceous) - includes Parascaniornis
- Family Hesperornithidae
- Genus Hesperornis (Late Cretaceous)
- Genus Parahesperornis (Late Cretaceous)
- Genus Canadaga (Late Cretaceous)
- Genus Coniornis (Late Cretaceous) - sometimes included in Hesperornis
- Family Enaliornithidae
Sometimes assigned to the Hesperornithes, but actual relationships unclear
- "cf. Parahesperornis" (Nemegt Late Cretaceous of Tsagaan Kushu, Mongolia) - hesperornithiform (hesperornithid)?
- Genus Neogaeornis (Late Cretaceous) - baptornithid, neornithine (gaviiform, procellariiform)?
- Genus Potamornis (Lance Late Cretaceous of Buck Creek, USA) - hesperornithid? baptornithid? enaliornithid?
Footnotes
- ↑ 1.0 1.1 Chinsamy et al. (1998)
- ↑ Discussed in Marsh (1880), Gregory, (1952).
- ↑ Discussed at length by Harris et al. (2006).
- ↑ E.g. by Cracraft (1982). That study is interesting example of how to arrive at an essentially phenetic - and utterly incorrect - analysis in spite of using cladistic methods.
- ↑ Based on a perceived similarity of the bony palate (Gingerich 1973).
- ↑ See Houde (1987).
- ↑ This is not a new theory; it was proposed by Stolpe as early as 1935. See also Bogdanovich (2003).
- ↑ Based on Marsh (1880), Feduccia (1996), Tokaryk, Cumbaa & Storer (1997), and Galton & Martin (2002) (via Haaramo, 2005), as well as Clarke (2004) and Sereno (2005)
- ↑ Mentioned but not discussed by Dyke et al. (2006): "This material probably pertains to a smaller [hesperornithine] taxon, an area for future work."
- ↑ A loon-sized hesperornithine (Longrich 2006).
References
- Bogdanovich, I.O. (2003): Морфологiчнi аспекти фiлогенii Hesperornithidae (Ornithurae, Aves) [Morphological Aspects of the Phylogeny of the Hesperornithidae (Ornithurae, Aves)]. Vestnik zoologii 37(6): 65–71. [Ukrainian with Russian and English abstract] PDF fulltext
- Chinsamy, A.; Martin, Larry D. & Dobson, P. (1998): Bone microstructure of the diving Hesperornis and the volant Ichthyornis from the Niobrara Chalk of western Kansas. Cretaceous Research 19(2): 225-235. doi:10.1006/cres.1997.0102 (HTML abstract)
- Clarke, J. A. (2004): Morphology, Phylogenetic Taxonomy, and Systematics of Ichthyornis and Apatornis (Avialae: Ornithurae). Bulletin of the American Museum of Natural History 286: 1-179 PDF fulltext
- Cracraft, Joel (1982): Phylogenetic relationships and monophyly of loons, grebes, and hesperornithiform birds, with comments on the early history of birds. Systematic Zoology 31: 35-56. doi:10.2307/2413412 (HTML abstract)
- Dyke, Gareth J.; Malakhov, Dmitry V. & Chiappe, Luis M. (2006): A re-analysis of the marine bird Asiahesperornis from northern Kazakhstan. Cretaceous Research 27(6): 947-953. doi:10.1016/j.cretres.2006.05.001 (HTML abstract)
- Everhart, Mike (2007): Oceans of Kansas: Hesperornis regalis Marsh 1872 - Toothed marine birds of the Late Cretaceous seas. Version of 2006-DEC-12. Retrieved 2007-AUG-23.
- Feduccia, Alan (1996): The origin and evolution of birds. Yale University Press, New Haven and London.
- Fürbringer, M. (1888): Untersuchungen zur Morphologie und Systematik der Vögel (2 vols). Von Holkema, Amsterdam.
- Galton, P. M. & Martin, Larry D. (2002): Enaliornis, an Early Cretaceous Hesperornithiform Bird from England, with Comments on Other Hesperornithiformes. 317-338. In: Chiappe, L. M. & Witmer, L. M. (eds.): Mesozoic Birds: Above the Heads of Dinosaurs. University of California Press, Berkeley, Los Angeles, London.
- Gingerich, P. D. (1973): Skull of Hesperornis and the early evolution of birds. Nature 243: 70-73. doi:10.1038/243070a0 (HTML abstract)
- Gregory, Joseph T. (1952): The Jaws of the Cretaceous Toothed Birds, Ichthyornis and Hesperornis. Condor 54(2): 73-88. PDF fulltext
- Haaramo, Mikko (2005): Mikko's Phylogeny Archive: †Hesperornithiformes. Version of 2005-11-15. Retrieved 2006-09-21.
- Harris, Matthew P.; Hasso, Sean M.; Ferguson, Mark W.J. & Fallon, John F. (2006): The development of archosaurian first-generation teeth in a chicken mutant. Current Biology 16(4): 371-377. PDF fulltext
- Houde, Peter (1987): Histological Evidence for the Systematic Position of Hesperornis (Odontornithes: Hesperornithiformes). Auk 104(1): 125-129 PDF fulltext
- Longrich, Nick (2006): An ornithurine bird from the Late Cretaceous of Alberta, Canada. Canadian Journal of Earth Sciences 43(1): 1-7. HTML abstract
- Marsh, Othniel Charles (1880): Odontornithes, a Monograph on the Extinct Toothed Birds of North America. Government Printing Office, Washington DC.
- Sereno, P. C. (2005): Hesperornithes. In: TaxonSearch: Stem Archosauria. Version 1.0, 2005 November 7. Retrieved 2006-09-21.
- Stolpe, M. (1935): Colymbus, Hesperornis, Podiceps:, ein Vergleich ihrer hinteren Extremität. J. Ornithol. 80(1): 161-247. [Article in German] doi:10.1007/BF01908745
- Tokaryk, T. T.; Cumbaa, S. L. & Storer, J. E. (1997): Early Late Cretaceous birds from the Saskatchewan, Canada: The oldest diverse avifauna known from North America. J. Vertebr. Paleontol.: 17(1): 172-176. HTML abstract
External links
|
||||||||||||||||||||||||||

|
||||||||||||||||||||
