HIV
| Human immunodeficiency virus | ||||||
|---|---|---|---|---|---|---|
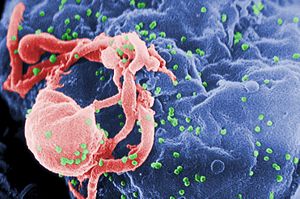 Scanning electron micrograph of HIV-1 (in green) budding from cultured lymphocyte. Multiple round bumps on cell surface represent sites of assembly and budding of virions.
|
||||||
| Virus classification | ||||||
|
||||||
| Species | ||||||
|
| International Statistical Classification of Diseases and Related Health Problems Codes Classification and external resources |
|
| ICD-10 | B20-B24 |
|---|---|
| ICD-9 | 042-044 |
Human immunodeficiency virus (HIV) is a lentivirus (a member of the retrovirus family) that can lead to acquired immunodeficiency syndrome (AIDS), a condition in humans in which the immune system begins to fail, leading to life-threatening opportunistic infections. Previous names for the virus include human T-lymphotropic virus-III (HTLV-III), lymphadenopathy-associated virus (LAV), and AIDS-associated retrovirus (ARV).[1][2]
Infection with HIV occurs by the transfer of blood, semen, vaginal fluid, pre-ejaculate, or breast milk. Within these bodily fluids, HIV is present as both free virus particles and virus within infected immune cells. The four major routes of transmission are unprotected sexual intercourse, contaminated needles, breast milk, and transmission from an infected mother to her baby at birth( Vertical transmission). Screening of blood products for HIV has largely eliminated transmission through blood transfusions or infected blood products in the developed world.
HIV infection in humans is now pandemic. As of January 2006, the Joint United Nations Programme on HIV/AIDS (UNAIDS) and the World Health Organization (WHO) estimate that AIDS has killed more than 25 million people since it was first recognized on December 1, 1981. It is estimated that about 0.6 percent of the world's population is infected with HIV.[3] In 2005 alone, AIDS claimed an estimated 2.4–3.3 million lives, of which more than 570,000 were children. A third of these deaths are occurring in sub-Saharan Africa, retarding economic growth and increasing poverty.[4] According to current estimates, HIV is set to infect 90 million people in Africa, resulting in a minimum estimate of 18 million orphans.[5] Antiretroviral treatment reduces both the mortality and the morbidity of HIV infection, but routine access to antiretroviral medication is not available in all countries.[6]
HIV primarily infects vital cells in the human immune system such as helper T cells (specifically CD4+ T cells), macrophages, and dendritic cells. HIV infection leads to low levels of CD4+ T cells through three main mechanisms: firstly, direct viral killing of infected cells; secondly, increased rates of apoptosis in infected cells; and thirdly, killing of infected CD4+ T cells by CD8 cytotoxic lymphocytes that recognize infected cells. When CD4+ T cell numbers decline below a critical level, cell-mediated immunity is lost, and the body becomes progressively more susceptible to opportunistic infections.
Eventually most HIV-infected individuals develop AIDS (Acquired Immunodeficiency Syndrome). These individuals mostly die from opportunistic infections or malignancies associated with the progressive failure of the immune system.[7] Without treatment, about 9 out of every 10 persons with HIV will progress to AIDS after 10-15 years. Many progress much sooner.[8] Treatment with anti-retrovirals increases the life expectancy of people infected with HIV. Even after HIV has progressed to diagnosable AIDS, the average survival time with antiretroviral therapy (as of 2005) is estimated to be more than 5 years.[9] Without antiretroviral therapy, death normally occurs within a year.[10] It is hoped that current and future treatments may allow HIV-infected individuals to achieve a life expectancy approaching that of the general public.
Contents |
Classification
HIV is a member of the genus Lentivirus,[11] part of the family of Retroviridae.[12] Lentiviruses have many common morphologies and biological properties. Many species are infected by lentiviruses, which are characteristically responsible for long-duration illnesses with a long incubation period.[13] Lentiviruses are transmitted as single-stranded, positive-sense, enveloped RNA viruses. Upon entry of the target cell, the viral RNA genome is converted to double-stranded DNA by a virally encoded reverse transcriptase that is present in the virus particle. This viral DNA is then integrated into the cellular DNA by a virally encoded integrase, along with host cellular co-factors,[14] so that the genome can be transcribed. Once the virus has infected the cell, two pathways are possible: either the virus becomes latent and the infected cell continues to function, or the virus becomes active and replicates, and a large number of virus particles are liberated that can then infect other cells.
There are two strains of HIV known to exist: HIV-1 and HIV-2. HIV-1 is the virus that was initially discovered and termed LAV. It is more virulent, relatively easily transmitted, and is the cause of the majority of HIV infections globally. HIV-2 is less transmittable and is largely confined to West Africa.[15]
| Species | Virulence | Transmittability | Prevalence | Purported origin |
|---|---|---|---|---|
| HIV-1 | High | High | Global | Common Chimpanzee |
| HIV-2 | Lower | Low | West Africa | Sooty Mangabey |
History
Origin
HIV is thought to have originated in non-human primates in sub-Saharan Africa and transferred to humans early in the 20th century.[16] The first paper recognizing a pattern of opportunistic infections was published on 4 June 1981.[17]
Two species of HIV infect humans: HIV-1 and HIV-2. Both species of the virus are believed to have originated in West-Central Africa and jumped species (zoonosis) from a non-human primate to humans. HIV-1 is thought to have originated in southern Cameroon after jumping from wild chimpanzees (Pan troglodytes troglodytes) to humans during the twentieth century.[18][19] It evolved from a Simian Immunodeficiency Virus (SIVcpz)[20] HIV-2, on the other hand, may have originated from the Sooty Mangabey (Cercocebus atys), an Old World monkey of Guinea-Bissau, Gabon, and Cameroon.[15]
New World Monkeys are an interesting exception to the transmission of HIV. Their immunity is believed to be caused by retrotransposition of the cyclophilin gene into an intron of TRIM5. The result is fusion gene that provides the owl monkey with resistance to HIV-1 infection. [21]
Early history
- See History of known cases and spread for early cases of HIV / AIDS
Discovery
Controversy surrounding the discovery of HIV was intense after French scientist Luc Montagnier and American researcher Robert Gallo both claimed to have discovered it, in 1983 and 1984 respectively.[22] In 1987 the dispute was initially settled on a political level with both teams receiving equal credit.[22] In 1991 a study confirmed that the samples in Gallo's laboratory had in fact originated in Montagnier's.[22] In 1994 the US Government conceded that the French should receive the lion's share of the credit.[23]
The Karolinska Institute awarded half of the 2008 Nobel Prize in Physiology or Medicine to Montagnier and his colleague Françoise Barré-Sinoussi 'for their discovery of "human immunodeficiency virus"'. The other half went to Harald zur Hausen for unrelated work on Human Papilloma Virus.[24] Gallo was reported to have said that it was "a disappointment" not to have been included, but that all three of the award's recipients deserved the honor. [25] The Karolinska Institute's press release stated "Soon after the discovery of the virus, several groups contributed to the definitive demonstration of HIV as the cause of acquired human immunodeficiency syndrome (AIDS)."[24]
Transmission
| Exposure Route | Estimated infections per 10,000 exposures to an infected source |
|||
|---|---|---|---|---|
| Blood Transfusion | 9,000[27] | |||
| Childbirth | 2,500[28] | |||
| Needle-sharing injection drug use | 67[29] | |||
| Percutaneous needle stick | 30[30] | |||
| Receptive anal intercourse* | 50[31][32] | |||
| Insertive anal intercourse* | 6.5[31][32] | |||
| Receptive penile-vaginal intercourse* | 10[31][32][33] | |||
| Insertive penile-vaginal intercourse* | 5[31][32] | |||
| Receptive oral intercourse*§ | 1[32] | |||
| Insertive oral intercourse*§ | 0.5[32] | |||
| * assuming no condom use § source refers to oral intercourse performed on a man |
||||
Three main transmission routes for HIV have been identified. HIV-2 is transmitted much less frequently by the mother-to-child and sexual route than HIV-1.
Sexual
The majority of HIV infections are acquired through unprotected sexual relations. Sexual transmission can occur when infected sexual secretions of one partner come into contact with the genital, oral, or rectal mucous membranes of another.
The correct and consistent use of latex condoms reduces the risk of sexual transmission of HIV by about 85%.[34] However, spermicide may actually increase the male to female transmission rate due to inflammation of the vagina.[35]
A meta-analysis of 27 observational studies conducted prior to 1999 in sub-Saharan Africa indicated that male circumcision reduces the risk of HIV infection.[36] However, a subsequent review indicated that the correlation between circumcision and HIV in these observational studies may have been due to confounding factors.[37] Later trials, in which uncircumcised men were randomly assigned to be medically circumcised in sterile conditions and given counseling and other men were not circumcised, have been conducted in South Africa,[38] Kenya[39] and Uganda[40] showing reductions in HIV transmission for heterosexual sex of 60 percent, 53 percent, and 51 percent respectively. As a result, a panel of experts convened by WHO and the UNAIDS Secretariat has "recommended that male circumcision now be recognized as an additional important intervention to reduce the risk of heterosexually acquired HIV infection in men."[41] Research is clarifying whether there is a historical relationship between rates of male circumcision and rates of HIV in differing social and cultural contexts.
On the other hand, some South African medical experts have expressed concern that the repeated use of unsterilized blades in the traditional circumcision of adolescent boys may actually be spreading HIV.[42]
Blood or blood product
In general if infected blood comes into contact with any open wound, HIV may be transmitted. This transmission route can account for infections in intravenous drug users, hemophiliacs and recipients of blood transfusions (though most transfusions are checked for HIV in the developed world) and blood products. It is also of concern for persons receiving medical care in regions where there is prevalent substandard hygiene in the use of injection equipment, such as the reuse of needles in Third World countries. Health care workers such as nurses, laboratory workers, and doctors have also been infected, although this occurs more rarely. People who give and receive tattoos, piercings, and scarification procedures can also be at risk of infection.
Since transmission of HIV by blood became known medical personnel are required to protect themselves from contact with blood by the use of universal precautions.
Mother-to-child
The transmission of the virus from the mother to the child can occur in utero during pregnancy and intrapartum at childbirth. In the absence of treatment, the transmission rate between the mother and child is around 25 percent.[28] However, where combination antiretroviral drug treatment and Cesarian section are available, this risk can be reduced to as low as one percent.[28]
Breast feeding also presents a risk of infection for the baby.
Other routes
HIV has been found at low concentrations in the saliva, tears and urine of infected individuals, but there are no recorded cases of infection by these secretions and the potential risk of transmission is negligible.[43]
Multiple infection
Unlike some other viruses, infection with HIV does not provide immunity against additional infections, particularly in the case of more genetically distant viruses. Both inter- and intra-clade multiple infections have been reported,[44] and even associated with more rapid disease progression.[45] Multiple infections are divided into two categories depending on the timing of the acquisition of the second strain. Coinfection refers to two strains that appear to have been acquired at the same time (or too close to distinguish). Reinfection (or superinfection) is infection with a second strain at a measurable time after the first. Both forms of dual infection have been reported for HIV in both acute and chronic infection around the world.[46][47][48][49]
Structure and genome

HIV is different in structure from other retroviruses. It is roughly spherical[50] with a diameter of about 120 nm, around 60 times smaller than a red blood cell, yet large for a virus.[51] It is composed of two copies of positive single-stranded RNA that codes for the virus's nine genes enclosed by a conical capsid composed of 2,000 copies of the viral protein p24.[52] The single-stranded RNA is tightly bound to nucleocapsid proteins, p7 and enzymes needed for the development of the virion such as reverse transcriptase, proteases, ribonuclease and integrase. A matrix composed of the viral protein p17 surrounds the capsid ensuring the integrity of the virion particle.[52] This is, in turn, surrounded by the viral envelope which is composed of two layers of fatty molecules called phospholipids taken from the membrane of a human cell when a newly formed virus particle buds from the cell. Embedded in the viral envelope are proteins from the host cell and about 70 copies of a complex HIV protein that protrudes through the surface of the virus particle.[52] This protein, known as Env, consists of a cap made of three molecules called glycoprotein (gp) 120, and a stem consisting of three gp41 molecules that anchor the structure into the viral envelope.[53] This glycoprotein complex enables the virus to attach to and fuse with target cells to initiate the infectious cycle.[53] Both these surface proteins, especially gp120, have been considered as targets of future treatments or vaccines against HIV.[54]
The RNA genome consists of at least 7 structural landmarks (LTR, TAR, RRE, PE, SLIP, CRS, INS) and nine genes (gag, pol, and env, tat, rev, nef, vif, vpr, vpu, and tev) encoding 19 proteins. Three of these genes, gag, pol, and env, contain information needed to make the structural proteins for new virus particles.[52] For example, env codes for a protein called gp160 that is broken down by a viral enzyme to form gp120 and gp41. The six remaining genes, tat, rev, nef, vif, vpr, and vpu (or vpx in the case of HIV-2), are regulatory genes for proteins that control the ability of HIV to infect cells, produce new copies of virus (replicate), or cause disease.[52] The two Tat proteins (p16 and p14) are transcriptional transactivators for the LTR promoter acting by binding the TAR RNA element. The Rev protein (p19) is involved in shuttling RNAs from the nucleus and the cytoplasm by binding to the RRE RNA element. The Vif protein (p23) prevents the action of APOBEC3G (a cell protein which deaminates DNA:RNA hybrids and/or interferes with the Pol protein). The Vpr protein (p14) arrests cell division at G2/M. The Nef protein (p27) downregulates CD4 (the major viral receptor), as well as the MHC class I and class II molecules.[55][56][57] Nef also interacts with SH3 domains. The Vpu protein (p16) influences the release of new virus particles from infected cells.[52] The ends of each strand of HIV RNA contain an RNA sequence called the long terminal repeat (LTR). Regions in the LTR act as switches to control production of new viruses and can be triggered by proteins from either HIV or the host cell. The Psi element is involved in viral genome packaging and recognized by Gag and Rev proteins. The SLIP element (TTTTTT) is involved in the frameshift in the Gag-Pol reading frame required to make functional Pol.[52]
Tropism
The term viral tropism refers to which cell types HIV infects. HIV can infect a variety of immune cells such as CD4+ T cells, macrophages, and microglial cells. HIV-1 entry to macrophages and CD4+ T cells is mediated through interaction of the virion envelope glycoproteins (gp120) with the CD4 molecule on the target cells and also with chemokine coreceptors.[53]
Macrophage (M-tropic) strains of HIV-1, or non-syncitia-inducing strains (NSI) use the β-chemokine receptor CCR5 for entry and are thus able to replicate in macrophages and CD4+ T cells.[58] This CCR5 coreceptor is used by almost all primary HIV-1 isolates regardless of viral genetic subtype. Indeed, macrophages play a key role in several critical aspects of HIV infection. They appear to be the first cells infected by HIV and perhaps the source of HIV production when CD4+ cells become depleted in the patient. Macrophages and microglial cells are the cells infected by HIV in the central nervous system. In tonsils and adenoids of HIV-infected patients, macrophages fuse into multinucleated giant cells that produce huge amounts of virus.
T-tropic isolates, or syncitia-inducing (SI) strains replicate in primary CD4+ T cells as well as in macrophages and use the α-chemokine receptor, CXCR4, for entry.[58][59][60] Dual-tropic HIV-1 strains are thought to be transitional strains of the HIV-1 virus and thus are able to use both CCR5 and CXCR4 as co-receptors for viral entry.
The α-chemokine SDF-1, a ligand for CXCR4, suppresses replication of T-tropic HIV-1 isolates. It does this by down-regulating the expression of CXCR4 on the surface of these cells. HIV that use only the CCR5 receptor are termed R5, those that only use CXCR4 are termed X4, and those that use both, X4R5. However, the use of coreceptor alone does not explain viral tropism, as not all R5 viruses are able to use CCR5 on macrophages for a productive infection[58] and HIV can also infect a subtype of myeloid dendritic cells,[61] which probably constitute a reservoir that maintains infection when CD4+ T cell numbers have declined to extremely low levels.
Some people are resistant to certain strains of HIV.[62] One example of how this occurs is people with the CCR5-Δ32 mutation; these people are resistant to infection with R5 virus as the mutation stops HIV from binding to this coreceptor, reducing its ability to infect target cells.
Sexual intercourse is the major mode of HIV transmission. Both X4 and R5 HIV are present in the seminal fluid which is passed from a male to his sexual partner. The virions can then infect numerous cellular targets and disseminate into the whole organism. However, a selection process leads to a predominant transmission of the R5 virus through this pathway.[63][64][65] How this selective process works is still under investigation, but one model is that spermatozoa may selectively carry R5 HIV as they possess both CCR3 and CCR5 but not CXCR4 on their surface[66] and that genital epithelial cells preferentially sequester X4 virus.[67] In patients infected with subtype B HIV-1, there is often a co-receptor switch in late-stage disease and T-tropic variants appear that can infect a variety of T cells through CXCR4.[68] These variants then replicate more aggressively with heightened virulence that causes rapid T cell depletion, immune system collapse, and opportunistic infections that mark the advent of AIDS.[69] Thus, during the course of infection, viral adaptation to the use of CXCR4 instead of CCR5 may be a key step in the progression to AIDS. A number of studies with subtype B-infected individuals have determined that between 40 and 50% of AIDS patients can harbour viruses of the SI, and presumably the X4, phenotype.[70][71]
Replication cycle

Entry to the cell
HIV enters macrophages and CD4+ T cells by the adsorption of glycoproteins on its surface to receptors on the target cell followed by fusion of the viral envelope with the cell membrane and the release of the HIV capsid into the cell.[72][73]
Entry to the cell begins through interaction of the trimeric envelope complex (gp160 spike) and both CD4 and a chemokine receptor (generally either CCR5 or CXCR4, but others are known to interact) on the cell surface.[72][73] gp120 binds to integrin α4β7 activating LFA-1 the central integrin involved in the establishment of virological synapses, which facilitate efficient cell-to-cell spreading of HIV-1.[74] The gp160 spike contains binding domains for both CD4 and chemokine receptors.[72][73] The first step in fusion involves the high-affinity attachment of the CD4 binding domains of gp120 to CD4. Once gp120 is bound with the CD4 protein, the envelope complex undergoes a structural change, exposing the chemokine binding domains of gp120 and allowing them to interact with the target chemokine receptor.[72][73] This allows for a more stable two-pronged attachment, which allows the N-terminal fusion peptide gp41 to penetrate the cell membrane.[72][73] Repeat sequences in gp41, HR1 and HR2 then interact, causing the collapse of the extracellular portion of gp41 into a hairpin. This loop structure brings the virus and cell membranes close together, allowing fusion of the membranes and subsequent entry of the viral capsid.[72][73]
Once HIV has bound to the target cell, the HIV RNA and various enzymes, including reverse transcriptase, integrase, ribonuclease and protease, are injected into the cell.[72] During the microtubule based transport to the nucleus, the viral single strand RNA genome is transcribed into double strand DNA, which is then integrated into a host chromosome.
HIV can infect dendritic cells (DCs) by this CD4-CCR5 route, but another route using mannose-specific C-type lectin receptors such as DC-SIGN can also be used.[75] DCs are one of the first cells encountered by the virus during sexual transmission. They are currently thought to play an important role by transmitting HIV to T cells once the virus has been captured in the mucosa by DCs.[75]
Replication and transcription
Once the viral capsid enters the cell, an enzyme called reverse transcriptase liberates the single-stranded (+)RNA from the attached viral proteins and copies it into a complementary DNA.[76] This process of reverse transcription is extremely error-prone and it is during this step that mutations may occur. Such mutations may cause drug resistance. The reverse transcriptase then makes a complementary DNA strand to form a double-stranded viral DNA intermediate (vDNA). This vDNA is then transported into the cell nucleus. The integration of the viral DNA into the host cell's genome is carried out by another viral enzyme called integrase.[76]
This integrated viral DNA may then lie dormant, in the latent stage of HIV infection.[76] To actively produce the virus, certain cellular transcription factors need to be present, the most important of which is NF-κB (NF kappa B), which is upregulated when T cells become activated.[77] This means that those cells most likely to be killed by HIV are those currently fighting infection.
In this replication process, the integrated provirus is copied to mRNA which is then spliced into smaller pieces. These small pieces produce the regulatory proteins Tat (which encourages new virus production) and Rev. As Rev accumulates it gradually starts to inhibit mRNA splicing.[78] At this stage, the structural proteins Gag and Env are produced from the full-length mRNA. The full-length RNA is actually the virus genome; it binds to the Gag protein and is packaged into new virus particles.
HIV-1 and HIV-2 appear to package their RNA differently; HIV-1 will bind to any appropriate RNA whereas HIV-2 will preferentially bind to the mRNA which was used to create the Gag protein itself. This may mean that HIV-1 is better able to mutate (HIV-1 infection progresses to AIDS faster than HIV-2 infection and is responsible for the majority of global infections).
Assembly and release
The final step of the viral cycle, assembly of new HIV-1 virons, begins at the plasma membrane of the host cell. The Env polyprotein (gp160) goes through the endoplasmic reticulum and is transported to the Golgi complex where it is cleaved by protease and processed into the two HIV envelope glycoproteins gp41 and gp120. These are transported to the plasma membrane of the host cell where gp41 anchors the gp120 to the membrane of the infected cell. The Gag (p55) and Gag-Pol (p160) polyproteins also associate with the inner surface of the plasma membrane along with the HIV genomic RNA as the forming virion begins to bud from the host cell. Maturation either occurs in the forming bud or in the immature virion after it buds from the host cell. During maturation, HIV proteases cleave the polyproteins into individual functional HIV proteins and enzymes. The various structural components then assemble to produce a mature HIV virion.[79] This cleavage step can be inhibited by protease inhibitors. The mature virus is then able to infect another cell.
Genetic variability
- Further information: Subtypes of HIV

HIV differs from many viruses in that it has very high genetic variability. This diversity is a result of its fast replication cycle, with the generation of 109 to 1010 virions every day, coupled with a high mutation rate of approximately 3 x 10-5 per nucleotide base per cycle of replication and recombinogenic properties of reverse transcriptase.[80] This complex scenario leads to the generation of many variants of HIV in a single infected patient in the course of one day.[80] This variability is compounded when a single cell is simultaneously infected by two or more different strains of HIV. When simultaneous infection occurs, the genome of progeny virions may be composed of RNA strands from two different strains. This hybrid virion then infects a new cell where it undergoes replication. As this happens, the reverse transcriptase, by jumping back and forth between the two different RNA templates, will generate a newly synthesized retroviral DNA sequence that is a recombinant between the two parental genomes.[80] This recombination is most obvious when it occurs between subtypes.[80]
The closely related simian immunodeficiency virus (SIV) exhibits a somewhat different behavior: in its natural hosts, African green monkeys and sooty mangabeys, the retrovirus is present in high levels in the blood, but evokes only a mild immune response,[81] does not cause the development of simian AIDS,[82] and does not undergo the extensive mutation and recombination typical of HIV.[83] By contrast, infection of heterologous hosts (rhesus or cynomologus macaques) with SIV results in the generation of genetic diversity that is on the same order as HIV in infected humans; these heterologous hosts also develop simian AIDS.[84] The relationship, if any, between genetic diversification, immune response, and disease progression is unknown.
Three groups of HIV-1 have been identified on the basis of differences in env: M, N, and O.[85] Group M is the most prevalent and is subdivided into eight subtypes (or clades), based on the whole genome, which are geographically distinct.[86] The most prevalent are subtypes B (found mainly in North America and Europe), A and D (found mainly in Africa), and C (found mainly in Africa and Asia); these subtypes form branches in the phylogenetic tree representing the lineage of the M group of HIV-1. Coinfection with distinct subtypes gives rise to circulating recombinant forms (CRFs). In 2000, the last year in which an analysis of global subtype prevalence was made, 47.2 percent of infections worldwide were of subtype C, 26.7 percent were of subtype A/CRF02_AG, 12.3 percent were of subtype B, 5.3 percent were of subtype D, 3.2 percent were of CRF_AE, and the remaining 5.3 percent were composed of other subtypes and CRFs.[87] Most HIV-1 research is focused on subtype B; few laboratories focus on the other subtypes.[88]
The genetic sequence of HIV-2 is only partially homologous to HIV-1 and more closely resembles that of SIV than HIV-1.
The clinical course of infection

Infection with HIV-1 is associated with a progressive decrease of the CD4+ T cell count and an increase in viral load. The stage of infection can be determined by measuring the patient's CD4+ T cell count, and the level of HIV in the blood.
HIV infection has basically four stages: incubation period, acute infection, latency stage and AIDS. The initial incubation period upon infection is asymptomatic and usually lasts between two and four weeks. The second stage, acute infection, which lasts an average of 28 days and can include symptoms such as fever, lymphadenopathy (swollen lymph nodes), pharyngitis (sore throat), rash, myalgia (muscle pain), malaise, and mouth and esophageal sores. The latency stage, which occurs third, shows few or no symptoms and can last anywhere from two weeks to twenty years and beyond. AIDS, the fourth and final stage of HIV infection shows as symptoms of various opportunistic infections.
Acute HIV infection
The initial infection with HIV generally occurs after transfer of body fluids from an infected person to an uninfected one. The first stage of infection, the primary, or acute infection, is a period of rapid viral replication that immediately follows the individual's exposure to HIV leading to an abundance of virus in the peripheral blood with levels of HIV commonly approaching several million viruses per mL.[89] This response is accompanied by a marked drop in the numbers of circulating CD4+ T cells. This acute viremia is associated in virtually all patients with the activation of CD8+ T cells, which kill HIV-infected cells, and subsequently with antibody production, or seroconversion. The CD8+ T cell response is thought to be important in controlling virus levels, which peak and then decline, as the CD4+ T cell counts rebound to around 800 cells per µL (the normal blood value is 1200 cells per µL ). A good CD8+ T cell response has been linked to slower disease progression and a better prognosis, though it does not eliminate the virus.[90] During this period (usually 2-4 weeks post-exposure) most individuals (80 to 90%) develop an influenza or mononucleosis-like illness called acute HIV infection, the most common symptoms of which may include fever, lymphadenopathy, pharyngitis, rash, myalgia, malaise, mouth and esophagal sores, and may also include, but less commonly, headache, nausea and vomiting, enlarged liver/spleen, weight loss, thrush, and neurological symptoms. Infected individuals may experience all, some, or none of these symptoms. The duration of symptoms varies, averaging 28 days and usually lasting at least a week.[91] Because of the nonspecific nature of these symptoms, they are often not recognized as signs of HIV infection. Even if patients go to their doctors or a hospital, they will often be misdiagnosed as having one of the more common infectious diseases with the same symptoms. Consequently, these primary symptoms are not used to diagnose HIV infection as they do not develop in all cases and because many are caused by other more common diseases. However, recognizing the syndrome can be important because the patient is much more infectious during this period.[92]
Latency stage
A strong immune defense reduces the number of viral particles in the blood stream, marking the start of the infection's clinical latency stage. Clinical latency can vary between two weeks and 20 years. During this early phase of infection, HIV is active within lymphoid organs, where large amounts of virus become trapped in the follicular dendritic cells (FDC) network.[93] The surrounding tissues that are rich in CD4+ T cells may also become infected, and viral particles accumulate both in infected cells and as free virus. Individuals who are in this phase are still infectious. During this time, CD4+ CD45RO+ T cells carry most of the proviral load.[94]
AIDS
- For more details on this topic, see AIDS Diagnosis, AIDS Symptoms and WHO Disease Staging System for HIV Infection and Disease
When CD4+ T cell numbers decline below a critical level, cell-mediated immunity is lost, and infections with a variety of opportunistic microbes appear. The first symptoms often include moderate and unexplained weight loss, recurring respiratory tract infections (such as sinusitis, bronchitis, otitis media, pharyngitis), prostatitis, skin rashes, and oral ulcerations. Common opportunistic infections and tumors, most of which are normally controlled by robust CD4+ T cell-mediated immunity then start to affect the patient. Typically, resistance is lost early on to oral Candida species and to Mycobacterium tuberculosis, which leads to an increased susceptibility to oral candidiasis (thrush) and tuberculosis. Later, reactivation of latent herpes viruses may cause worsening recurrences of herpes simplex eruptions, shingles, Epstein-Barr virus-induced B-cell lymphomas, or Kaposi's sarcoma, a tumor of endothelial cells that occurs when HIV proteins such as Tat interact with Human Herpesvirus-8. Pneumonia caused by the fungus Pneumocystis jirovecii is common and often fatal. In the final stages of AIDS, infection with cytomegalovirus (another herpes virus) or Mycobacterium avium complex is more prominent. Not all patients with AIDS get all these infections or tumors, and there are other tumors and infections that are less prominent but still significant.
HIV test
Many HIV-positive people are unaware that they are infected with the virus.[95] For example, less than 1% of the sexually active urban population in Africa have been tested and this proportion is even lower in rural populations.[95] Furthermore, only 0.5% of pregnant women attending urban health facilities are counselled, tested or receive their test results.[95] Again, this proportion is even lower in rural health facilities.[95] Since donors may therefore be unaware of their infection, donor blood and blood products used in medicine and medical research are routinely screened for HIV.[96]
HIV-1 testing consists of initial screening with an enzyme-linked immunosorbent assay (ELISA) to detect antibodies to HIV-1. Specimens with a nonreactive result from the initial ELISA are considered HIV-negative unless new exposure to an infected partner or partner of unknown HIV status has occurred. Specimens with a reactive ELISA result are retested in duplicate.[97] If the result of either duplicate test is reactive, the specimen is reported as repeatedly reactive and undergoes confirmatory testing with a more specific supplemental test (e.g., Western blot or, less commonly, an immunofluorescence assay (IFA)). Only specimens that are repeatedly reactive by ELISA and positive by IFA or reactive by Western blot are considered HIV-positive and indicative of HIV infection. Specimens that are repeatedly ELISA-reactive occasionally provide an indeterminate Western blot result, which may be either an incomplete antibody response to HIV in an infected person, or nonspecific reactions in an uninfected person.[98] Although IFA can be used to confirm infection in these ambiguous cases, this assay is not widely used. Generally, a second specimen should be collected more than a month later and retested for persons with indeterminate Western blot results. Although much less commonly available, nucleic acid testing (e.g., viral RNA or proviral DNA amplification method) can also help diagnosis in certain situations.[97] In addition, a few tested specimens might provide inconclusive results because of a low quantity specimen. In these situations, a second specimen is collected and tested for HIV infection.
Treatment
- See also Antiretroviral drug

There is currently no vaccine or cure for HIV or AIDS.[99] The only known method of prevention is avoiding exposure to the virus. However, a course of antiretroviral treatment administered immediately after exposure, referred to as post-exposure prophylaxis, is believed to reduce the risk of infection if begun as quickly as possible.[100] Current treatment for HIV infection consists of highly active antiretroviral therapy, or HAART.[101] This has been highly beneficial to many HIV-infected individuals since its introduction in 1996, when the protease inhibitor-based HAART initially became available.[102] Current HAART options are combinations (or "cocktails") consisting of at least three drugs belonging to at least two types, or "classes," of antiretroviral agents. Typically, these classes are two nucleoside analogue reverse transcriptase inhibitors (NARTIs or NRTIs) plus either a protease inhibitor or a non-nucleoside reverse transcriptase inhibitor (NNRTI). New classes of drugs such as Entry Inhibitors provide treatment options for patients who are infected with viruses already resistant to common therapies, although they are not widely available and not typically accessible in resource-limited settings. Because AIDS progression in children is more rapid and less predictable than in adults, particularly in young infants, more aggressive treatment is recommended for children than adults.[103] In developed countries where HAART is available, doctors assess their patients thoroughly: measuring the viral load, how fast CD4 declines, and patient readiness. They then decide when to recommend starting treatment.[104]
HAART neither cures the patient nor does it uniformly remove all symptoms; high levels of HIV-1, often HAART resistant, return if treatment is stopped.[105][106] Moreover, it would take more than a lifetime for HIV infection to be cleared using HAART.[107] Despite this, many HIV-infected individuals have experienced remarkable improvements in their general health and quality of life, which has led to a large reduction in HIV-associated morbidity and mortality in the developed world.[102][108][109] One study suggests the average life expectancy of an HIV infected individual is 32 years from the time of infection if treatment is started when the CD4 count is 350/µL.[110] In the absence of HAART, progression from HIV infection to AIDS has been observed to occur at a median of between nine to ten years and the median survival time after developing AIDS is only 9.2 months.[10] However, HAART sometimes achieves far less than optimal results, in some circumstances being effective in less than fifty percent of patients. This is due to a variety of reasons such as medication intolerance/side effects, prior ineffective antiretroviral therapy and infection with a drug-resistant strain of HIV. However, non-adherence and non-persistence with antiretroviral therapy is the major reason most individuals fail to benefit from HAART.[111] The reasons for non-adherence and non-persistence with HAART are varied and overlapping. Major psychosocial issues, such as poor access to medical care, inadequate social supports, psychiatric disease and drug abuse contribute to non-adherence. The complexity of these HAART regimens, whether due to pill number, dosing frequency, meal restrictions or other issues along with side effects that create intentional non-adherence also contribute to this problem.[112][113][114] The side effects include lipodystrophy, dyslipidemia, insulin resistance, an increase in cardiovascular risks and birth defects.[115][116]
The timing for starting HIV treatment is still debated. There is no question that treatment should be started before the patient's CD4 count falls below 200, and most national guidelines say to start treatment once the CD4 count falls below 350; but there is some evidence from cohort studies that treatment should be started before the CD4 count falls below 350.[117][108] In those countries where CD4 counts are not available, patients with WHO stage III or IV disease[118] should be offered treatment.
Anti-retroviral drugs are expensive, and the majority of the world's infected individuals do not have access to medications and treatments for HIV and AIDS.[119] Research to improve current treatments includes decreasing side effects of current drugs, further simplifying drug regimens to improve adherence, and determining the best sequence of regimens to manage drug resistance. Unfortunately, only a vaccine is thought to be able to halt the pandemic. This is because a vaccine would cost less, thus being affordable for developing countries, and would not require daily treatment.[119] However, after over 20 years of research, HIV-1 remains a difficult target for a vaccine.[119]
Treatments in development
Promising new treatments include Cre recombinase[120] and the enzyme Tre recombinase,[121] both of which are able to remove HIV from an infected cell. These enzymes promise a treatment in which a patient's stem cells are extracted, cured, and reinjected to promulgate the enzyme into the body. The carried enzyme then finds and removes the virus.
In 2008, Dr. Gero Hütter announced that an HIV-positive leukemia patient had been "functionally cured" following a bone marrow transplant from a compatible donor who possessed the CCR5-Δ32 mutation (which confers resistance to HIV).[122] After 600 days without antiretroviral drug treatment, no HIV was detectable in the patient's blood or tissues. The mortality risk associated with bone marrow transplants may contraindicate the use of this treatment for HIV-positive individuals without leukemia or lymphoma.[122]
Epidemiology

UNAIDS and the WHO estimate that AIDS has killed more than 25 million people since it was first recognized in 1981, making it one of the most destructive pandemics in recorded history. Despite recent improved access to antiretroviral treatment and care in many regions of the world, the AIDS pandemic claimed an estimated 2.8 million (between 2.4 and 3.3 million) lives in 2005 of which more than half a million (570,000) were children.[3]
In 2007, between 30.6 and 36.1 million people were believed to live with HIV, and it killed an estimated 2.1 million people that year, including 330,000 children; there were 2.5 million new infections.[123]
Sub-Saharan Africa remains by far the worst-affected region, with an estimated 21.6 to 27.4 million people currently living with HIV. Two million [1.5–3.0 million] of them are children younger than 15 years of age. More than 64% of all people living with HIV are in sub-Saharan Africa, as are more than three quarters of all women living with HIV. In 2005, there were 12.0 million [10.6–13.6 million] AIDS orphans living in sub-Saharan Africa 2005.[3] South & South East Asia are second-worst affected with 15% of the total. AIDS accounts for the deaths of 500,000 children in this region. South Africa has the largest number of HIV patients in the world followed by Nigeria.[124] India has an estimated 2.5 million infections (0.23% of population), making India the country with the third largest population of HIV patients. In the 35 African nations with the highest prevalence, average life expectancy is 48.3 years—6.5 years less than it would be without the disease.[125]
The latest evaluation report of the World Bank's Operations Evaluation Department assesses the development effectiveness of the World Bank's country-level HIV/AIDS assistance defined as policy dialogue, analytic work, and lending with the explicit objective of reducing the scope or impact of the AIDS epidemic.[126] This is the first comprehensive evaluation of the World Bank's HIV/AIDS support to countries, from the beginning of the epidemic through mid-2004. Because the Bank aims to assist in implementation of national government programmes, their experience provides important insights on how national AIDS programmes can be made more effective.
The development of HAART as effective therapy for HIV infection and AIDS has substantially reduced the death rate from this disease in those areas where these drugs are widely available. This has created the misperception that the disease has vanished. In fact, as the life expectancy of persons with AIDS has increased in countries where HAART is widely used, the number of persons living with AIDS has increased substantially. In the United States, the number of persons with AIDS increased from about 35,000 in 1988 to over 220,000 in 1996 and 312,000 in 2002[127]
In Africa, the number of MTCT and the prevalence of AIDS is beginning to reverse decades of steady progress in child survival. Countries such as Uganda are attempting to curb the MTCT epidemic by offering VCT (voluntary counselling and testing), PMTCT (prevention of mother-to-child transmission) and ANC (ante-natal care) services, which include the distribution of antiretroviral therapy.
AIDS denialism
Some individuals, including some scientists who are not recognized experts on HIV, question the connection between HIV and AIDS. [128] Some question the procedures used by Montagnier's group in 1983, as well as other groups subsequently, to prove the existence of HIV. [129] Others question the validity of current testing and treatment methods. These claims have been examined and rejected as having no validity,[130] although they have had a political impact, particularly in South Africa, where governmental acceptance of AIDS denialism has been blamed for an ineffective response to that country's AIDS epidemic.[131][132][133]
References
- ↑ Rick Sowadsky (1999-02-24). "What is HTLV-III?". Retrieved on 2008-04-03.
- ↑ Coffin, J., Haase, A., Levy, J. A., Montagnier, L., Oroszlan, S., Teich, N., Temin, H., Toyoshima, K., Varmus, H., Vogt, P. and Weiss, R. A. (1986). "What to call the AIDS virus?". Nature 321 (6065): 10. doi:. PMID 3010128.
- ↑ 3.0 3.1 3.2 Joint United Nations Programme on HIV/AIDS (2006). "Overview of the global AIDS epidemic" (PDF format). 2006 Report on the global AIDS epidemic.
- ↑ Greener, R. (2002). "AIDS and macroeconomic impact". in S, Forsyth (ed.). State of The Art: AIDS and Economics. IAEN. pp. 49–55.
- ↑ Joint United Nations Programme on HIV/AIDS. "AIDS epidemic update, 2005" (PDF format). Retrieved on 2006-02-28.
- ↑ Palella, F. J. Jr, Delaney, K. M., Moorman, A. C., Loveless, M. O., Fuhrer, J., Satten, G. A., Aschman and D. J., Holmberg, S. D. (1998). "Declining morbidity and mortality among patients with advanced human immunodeficiency virus infection. HIV Outpatient Study Investigators". N. Engl. J. Med 338 (13): 853–860. doi:. PMID 9516219.
- ↑ Lawn SD (2004). "AIDS in Africa: the impact of coinfections on the pathogenesis of HIV-1 infection". J. Infect. Dis. 48 (1): 1–12. PMID 14667787.
- ↑ Buchbinder SP, Katz MH, Hessol NA, O'Malley PM, Holmberg SD. (1994). "Long-term HIV-1 infection without immunologic progression". AIDS 8 (8): 1123–8. doi:. PMID 7986410.
- ↑ Schneider MF, Gange SJ, Williams CM, Anastos K, Greenblatt RM, Kingsley L, Detels R, Munoz A (2005). "Patterns of the hazard of death after AIDS through the evolution of antiretroviral therapy: 1984–2004". AIDS 19 (17): 2009–18. PMID 16260908.
- ↑ 10.0 10.1 Morgan D, Mahe C, Mayanja B, Okongo JM, Lubega R, Whitworth JA (2002). "HIV-1 infection in rural Africa: is there a difference in median time to AIDS and survival compared with that in industrialized countries?". AIDS 16 (4): 597–632. PMID 11873003.
- ↑ International Committee on Taxonomy of Viruses. "61.0.6. Lentivirus". National Institutes of Health. Retrieved on 2006-02-28.
- ↑ International Committee on Taxonomy of Viruses. "61. Retroviridae". National Institutes of Health. Retrieved on 2006-02-28.
- ↑ Lévy, J. A. (1993). "HIV pathogenesis and long-term survival". AIDS 7 (11): 1401–10. doi:. PMID 8280406.
- ↑ Smith, Johanna A.; Daniel, René (Division of Infectious Diseases, Center for Human Virology, Thomas Jefferson University, Philadelphia) (2006). "Following the path of the virus: the exploitation of host DNA repair mechanisms by retroviruses". ACS Chem Biol 1 (4): 217–26. doi:. PMID 17163676.
- ↑ 15.0 15.1 Reeves, J. D. and Doms, R. W (2002). "Human Immunodeficiency Virus Type 2". J. Gen. Virol. 83 (Pt 6): 1253–65. PMID 12029140.
- ↑ "Worobey et al : : Nature".
- ↑ "Pneumocystis Pneumonia -- Los Angeles". Retrieved on 2008-05-05.
- ↑ Gao, F., Bailes, E., Robertson, D. L., Chen, Y., Rodenburg, C. M., Michael, S. F., Cummins, L. B., Arthur, L. O., Peeters, M., Shaw, G. M., Sharp, P. M., and Hahn, B. H. (1999). "Origin of HIV-1 in the Chimpanzee Pan troglodytes troglodytes". Nature 397 (6718): 436–441. doi:. PMID 9989410.
- ↑ Keele, B. F., van Heuverswyn, F., Li, Y. Y., Bailes, E., Takehisa, J., Santiago, M. L., Bibollet-Ruche, F., Chen, Y., Wain, L. V., Liegois, F., Loul, S., Mpoudi Ngole, E., Bienvenue, Y., Delaporte, E., Brookfield, J. F. Y., Sharp, P. M., Shaw, G. M., Peeters, M., and Hahn, B. H. (2006). "Chimpanzee Reservoirs of Pandemic and Nonpandemic HIV-1". Science Online 2006-05-25: 523. doi:. PMID 16728595. http://www.sciencemag.org/cgi/content/abstract/1126531.
- ↑ http://www.nature.com/nature/journal/v397/n6718/abs/397436a0_fs.html
- ↑ Goodier, J., and Kazazian, H. (2008). "Retrotransposons Revisited: The Restraint and Rehabilitation of Parasites". Cell 135 (1): 23–35. doi:. http://www.sciencedirect.com/science/article/B6WSN-4TK9636-8/2/2af3e1e709ed15939505cf676ce4957c. Retrieved on 2008-10-10.
- ↑ 22.0 22.1 22.2 Brown, Phyllida (25 May 1991). "The strains of the HIV war". New Scientist. http://www.newscientist.com/channel/health/mg13017703.800-the-strains-of-the-hiv-war.html.
- ↑ "Nobel Prize awarded for AIDS, cervical cancer research -". Los Angeles Times. Retrieved on 2008-10-06.
- ↑ 24.0 24.1 Karolinska Institute (2008-10-06). "The Nobel Prize in Physiology or Medicine 2008". Press release. Retrieved on 2008-10-07.
- ↑ Ritter, K and Moore, M (October 6, 2008). "3 European scientists share Nobel medicine prize", Associated Press. Retrieved on 2008-10-06.
- ↑ Smith DK, Grohskopf LA, Black RJ, et al (2005). "Antiretroviral Postexposure Prophylaxis After Sexual, Injection-Drug Use, or Other Nonoccupational Exposure to HIV in the United States". MMWR 54 (RR02): 1–20. http://www.cdc.gov/mmwr/preview/mmwrhtml/rr5402a1.htm#tab1.
- ↑ Donegan E, Stuart M, Niland JC, et al (1990). "Infection with human immunodeficiency virus type 1 (HIV-1) among recipients of antibody-positive blood donations". Ann. Intern. Med. 113 (10): 733–739. PMID 2240875.
- ↑ 28.0 28.1 28.2 Coovadia H (2004). "Antiretroviral agents—how best to protect infants from HIV and save their mothers from AIDS". N. Engl. J. Med. 351 (3): 289–292. PMID 15247337.
- ↑ Kaplan EH, Heimer R (1995). "HIV incidence among New Haven needle exchange participants: updated estimates from syringe tracking and testing data". J. Acquir. Immune Defic. Syndr. Hum. Retrovirol. 10 (2): 175–176. PMID 7552482.
- ↑ Bell DM (1997). "Occupational risk of human immunodeficiency virus infection in healthcare workers: an overview.". Am. J. Med. 102 (5B): 9–15. PMID 9845490.
- ↑ 31.0 31.1 31.2 31.3 European Study Group on Heterosexual Transmission of HIV (1992). "Comparison of female to male and male to female transmission of HIV in 563 stable couples". BMJ. 304 (6830): 809–813. PMID 1392708.
- ↑ 32.0 32.1 32.2 32.3 32.4 32.5 Varghese B, Maher JE, Peterman TA, Branson BM,Steketee RW (2002). "Reducing the risk of sexual HIV transmission: quantifying the per-act risk for HIV on the basis of choice of partner, sex act, and condom use". Sex. Transm. Dis. 29 (1): 38–43. PMID 11773877.
- ↑ Leynaert B, Downs AM, de Vincenzi I (1998). "Heterosexual transmission of human immunodeficiency virus: variability of infectivity throughout the course of infection. European Study Group on Heterosexual Transmission of HIV". Am. J. Epidemiol. 148 (1): 88–96. PMID 9663408.
- ↑ National Institute of Allergy and Infectious Diseases; National Institutes of Health, Department of Health and Human Services (2001-07-20). "Workshop Summary: Scientific Evidence on Condom Effectiveness for Sexually Transmitted Disease (STD) Prevention" (PDF).: pp.13-15. Retrieved on 2007-04-07.
- ↑ "Should spermicides be used with condoms?". Condom Brochure, FDA OSHI HIV STDs. Retrieved on 2006-10-23.
- ↑ Weiss, H.A.; M.A. Quigley, R.J. Hayes (2000). "Male circumcision and risk of HIV infection in sub-Saharan Africa: A systematic review and meta-analysis". AIDS 14 (15): 2361–70. doi:. PMID 11089625.
- ↑ Siegfried, N., Muller, M., Deeks, J., Volmink, J., Egger, M., Low, N., Walker, S. and Williamson, P. (2005). "HIV and male circumcision--a systematic review with assessment of the quality of studies". Lancet Infect. Dis. 5 (3): 165–73. PMID 15766651.
- ↑ Williams BG, Lloyd-Smith JO, Gouws E, Hankins C, Getz WM, Hargrove J, de Zoysa I, Dye C, Auvert B. (2006). "The Potential Impact of Male Circumcision on HIV in Sub-Saharan Africa.". PLoS Med 3 (7): e262. PMID 16822094.
- ↑ Bailey RC, Moses S, Parker CB, et al (2007). "Male circumcision for HIV prevention in young men in Kisumu, Kenya: a randomised controlled trial". Lancet 369 (9562): 643–56. doi:. PMID 17321310.
- ↑ Gray RH et al (24 February 2007). "Male circumcision for HIV prevention in men in Rakai, Uganda: a randomised trial". Lancet 369 (9562): 657–66. doi:. PMID 17321311.
- ↑ WHO (2007). "WHO and UNAIDS announce recommendations from expert consultation on male circumcision for HIV prevention". WHO.int. Retrieved on 2007-07-13.
- ↑ Various (2005). "Repeated Use of Unsterilized Blades in Ritual Circumcision Might Contribute to HIV Spread in S. Africa, Doctors Say". Kaisernetwork.org. Retrieved on 2006-03-28.
- ↑ Lifson AR (1988). "Do alternate modes for transmission of human immunodeficiency virus exist? A review". JAMA 259 (9): 1353–6. PMID 2963151.
- ↑ Smith D, Richman D, Little S (2005). "HIV Superinfection". Journal of Infectious Diseases 192: 438–44. doi:. PMID 15995957.
- ↑ Gottlieb, et al (2004). "Dual HIV-1 infection associated with rapid disease progression". Lancet 363 (9049): 619–22. doi:. PMID 14987889.
- ↑ Smith et al (2004). "Incidence of HIV superinfection following primary infection". JAMA 292 (10): 1177–8. doi:. PMID 15353529. http://jama.ama-assn.org/cgi/content/full/292/10/1177.
- ↑ Chohan B, Lavreys L, Rainwater SM, Overbaugh J (Aug 2005). "Evidence for frequent reinfection with human immunodeficiency virus type 1 of a different subtype". J. Virol. 79 (16): 10701–8. doi:. PMID 16051862.
- ↑ Piantadosi A, Chohan B, Chohan V, McClelland RS, Overbaugh J (Nov 2007). "Chronic HIV-1 infection frequently fails to protect against superinfection". PLoS Pathog. 3 (11): e177. doi:. PMID 18020705.
- ↑ Hu DJ, Subbarao S, Vanichseni S, et al (Feb 2005). "Frequency of HIV-1 dual subtype infections, including intersubtype superinfections, among injection drug users in Bangkok, Thailand". AIDS 19 (3): 303–8. PMID 15718841. http://meta.wkhealth.com/pt/pt-core/template-journal/lwwgateway/media/landingpage.htm?an=00002030-200502180-00009.
- ↑ McGovern SL, Caselli E, Grigorieff N, Shoichet BK (2002). "A common mechanism underlying promiscuous inhibitors from virtual and high-throughput screening". J Med Chem 45 (8): 1712–22. PMID 11931626.
- ↑ Compared with overview in: Fisher, Bruce; Harvey, Richard P.; Champe, Pamela C.. Lippincott's Illustrated Reviews: Microbiology (Lippincott's Illustrated Reviews Series). Hagerstown, MD: Lippincott Williams & Wilkins. ISBN 0-7817-8215-5. Page 3
- ↑ 52.0 52.1 52.2 52.3 52.4 52.5 52.6 Various (2008) (PDF format). HIV Sequence Compendium 2008 Introduction. http://www.hiv.lanl.gov/content/sequence/HIV/COMPENDIUM/2008/frontmatter.pdf.
- ↑ 53.0 53.1 53.2 Chan, DC., Fass, D., Berger, JM., Kim, PS. (1997). "Core Structure of gp41 from the HIV Envelope Glycoprotein" (pdf). Cell 89: 263–73. PMID 9108481. http://www.its.caltech.edu/~chanlab/PDFs/Chan_Cell_1997.pdf.
- ↑ National Institute of Health (June 17, 1998). "Crystal Structure of Key HIV Protein Reveals New Prevention, Treatment Targets". Retrieved on 2006-09-14.
- ↑ Garcia JV, Miller AD (April 1991). "Serine phosphorylation-independent downregulation of cell-surface CD4 by nef". Nature 350 (6318): 508–11. doi:. PMID 2014052.
- ↑ Schwartz O, Maréchal V, Le Gall S, Lemonnier F, Heard JM (March 1996). "Endocytosis of major histocompatibility complex class I molecules is induced by the HIV-1 Nef protein". Nat. Med. 2 (3): 338–42. PMID 8612235.
- ↑ Stumptner-Cuvelette P, Morchoisne S, Dugast M, et al (October 2001). "HIV-1 Nef impairs MHC class II antigen presentation and surface expression". Proc. Natl. Acad. Sci. U.S.A. 98 (21): 12144–9. doi:. PMID 11593029.
- ↑ 58.0 58.1 58.2 Coakley, E., Petropoulos, C. J. and Whitcomb, J. M. (2005). "Assessing ch vbgemokine co-receptor usage in HIV". Curr. Opin. Infect. Dis. 18 (1): 9–15. PMID 15647694.
- ↑ Deng H, Liu R, Ellmeier W, Choe S, Unutmaz D, Burkhart M, Di Marzio P, Marmon S, Sutton RE, Hill CM, Davis CB, Peiper SC, Schall TJ, Littman DR, Landau NR. (1996). "Identification of a major co-receptor for primary isolates of HIV-1". Nature 381 (6584): 661–6. doi:. PMID 8649511.
- ↑ Feng Y, Broder CC, Kennedy PE, Berger EA. (1996). "HIV-1 entry cofactor: functional cDNA cloning of a seven-transmembrane, G protein-coupled receptor". Science 272 (5263): 872–7. doi:. PMID 8629022.
- ↑ Knight, S. C., Macatonia, S. E. and Patterson, S. (1990). "HIV I infection of dendritic cells". Int. Rev. Immunol. 6 (2-3): 163–75. doi:. PMID 2152500.
- ↑ Tang, J. and Kaslow, R. A. (2003). "The impact of host genetics on HIV infection and disease progression in the era of highly active antiretroviral therapy". AIDS 17 (Suppl 4): S51–S60. PMID 15080180.
- ↑ Zhu T, Mo H, Wang N, Nam DS, Cao Y, Koup RA, Ho DD. (1993). "Genotypic and phenotypic characterization of HIV-1 patients with primary infection". Science 261 (5125): 1179–81. doi:. PMID 8356453.
- ↑ van’t Wout AB, Kootstra NA, Mulder-Kampinga GA, Albrecht-van Lent N, Scherpbier HJ, Veenstra J, Boer K, Coutinho RA, Miedema F, Schuitemaker H. (1994). "Macrophage-tropic variants initiate human immunodeficiency virus type 1 infection after sexual, parenteral, and vertical transmission". J Clin Invest 94 (5): 2060–7. doi:. PMID 7962552.
- ↑ Zhu T, Wang N, Carr A, Nam DS, Moor-Jankowski R, Cooper DA, Ho DD. (1996). "Genetic characterization of human immunodeficiency virus type 1 in blood and genital secretions: evidence for viral compartmentalization and selection during sexual transmission". J Virol 70 (5): 3098–107. PMID 8627789.
- ↑ Muciaccia B, Padula F, Vicini E, Gandini L, Lenzi A, Stefanini M. (2005). "Beta-chemokine receptors 5 and 3 are expressed on the head region of human spermatozoon". Faseb J 19 (14): 2048–50. PMID 16174786.
- ↑ Berlier W, Bourlet T, Lawrence P, Hamzeh H, Lambert C, Genin C, Verrier B, Dieu-Nosjean MC, Pozzetto B, Delezay O. (2005). "Selective sequestration of X4 isolates by human genital epithelial cells: Implication for virus tropism selection process during sexual transmission of HIV". J Med Virol. 77 (4): 465–74. doi:. PMID 16254974.
- ↑ Clevestig P, Maljkovic I, Casper C, Carlenor E, Lindgren S, Naver L, Bohlin AB, Fenyo EM, Leitner T, Ehrnst A. (2005). "The X4 phenotype of HIV type 1 evolves from R5 in two children of mothers, carrying X4, and is not linked to transmission". AIDS Res Hum Retroviruses 5 (21): 371–8. doi:. PMID 15929699.
- ↑ Moore JP. (1997). "Coreceptors: implications for HIV pathogenesis and therapy". Science 276 (5309): 51–2. doi:. PMID 9122710.
- ↑ Karlsson A, Parsmyr K, Aperia K, Sandstrom E, Fenyo EM, Albert J. (1994). "MT-2 cell tropism of human immunodeficiency virus type 1 isolates as a marker for response to treatment and development of drug resistance". J Infect Dis. 170 (6): 1367–75. PMID 7995974.
- ↑ Koot M, van 't Wout AB, Kootstra NA, de Goede RE, Tersmette M, Schuitemaker H. (1996). "Relation between changes in cellular load, evolution of viral phenotype, and the clonal composition of virus populations in the course of human immunodeficiency virus type 1 infection". J Infect Dis. 173 (2): 349–54. PMID 8568295.
- ↑ 72.0 72.1 72.2 72.3 72.4 72.5 72.6 Chan D, Kim P (1998). "HIV entry and its inhibition". Cell 93 (5): 681–4. doi:. PMID 9630213.
- ↑ 73.0 73.1 73.2 73.3 73.4 73.5 Wyatt R, Sodroski J (1998). "The HIV-1 envelope glycoproteins: fusogens, antigens, and immunogens". Science 280 (5371): 1884–8. doi:. PMID 9632381.
- ↑ Arthos J, Cicala C, Martinelli E, Macleod K, Van Ryk D, Wei D, Xiao Z, Veenstra TD, Conrad TP, Lempicki RA, McLaughlin S, Pascuccio M, Gopaul R, McNally J, Cruz CC, Censoplano N, Chung E, Reitano KN, Kottilil S, Goode DJ, Fauci AS. (2008). "HIV-1 envelope protein binds to and signals through integrin alpha(4)beta(7), the gut mucosal homing receptor for peripheral T cells". Nature Immunol. In Press: 301. doi:. PMID 18264102.
- ↑ 75.0 75.1 Pope M, Haase A (2003). "Transmission, acute HIV-1 infection and the quest for strategies to prevent infection". Nat Med 9 (7): 847–52. doi:. PMID 12835704.
- ↑ 76.0 76.1 76.2 Zheng, Y. H., Lovsin, N. and Peterlin, B. M. (2005). "Newly identified host factors modulate HIV replication". Immunol. Lett. 97 (2): 225–34. doi:. PMID 15752562.
- ↑ Hiscott J, Kwon H, Genin P. (2001). "Hostile takeovers: viral appropriation of the NF-kappaB pathway". J Clin Invest. 107 (2): 143–151. doi:. PMID 11160127.
- ↑ Pollard, V. W. and Malim, M. H. (1998). "The HIV-1 Rev protein". Annu. Rev. Microbiol. 52: 491–532. doi:. PMID 9891806.
- ↑ Gelderblom, H. R (1997). "Fine structure of HIV and SIV". in Los Alamos National Laboratory (ed.) (PDF format). HIV Sequence Compendium. Los Alamos, New Mexico: Los Alamos National Laboratory. pp. 31–44.
- ↑ 80.0 80.1 80.2 80.3 Robertson DL, Hahn BH, Sharp PM. (1995). "Recombination in AIDS viruses". J Mol Evol. 40 (3): 249–59. doi:. PMID 7723052.
- ↑ Holzammer S, Holznagel E, Kaul A, Kurth R, Norley S (2001). "High virus loads in naturally and experimentally SIVagm-infected African green monkeys". Virology 283 (2): 324–31. doi:. PMID 11336557.
- ↑ Kurth, R. and Norley, S. (1996) Why don't the natural hosts of SIV develop simian AIDS?, J. NIH Res. 8, 33-37.
- ↑ Baier M, Dittmar MT, Cichutek K, Kurth R (1991). "Development of vivo of genetic variability of simian immunodeficiency virus". Proc. Natl. Acad. Sci. U.S.A. 88 (18): 8126–30. PMID 1896460.
- ↑ Daniel MD, King NW, Letvin NL, Hunt RD, Sehgal PK, Desrosiers RC (1984). "A new type D retrovirus isolated from macaques with an immunodeficiency syndrome". Science 223 (4636): 602–5. doi:. PMID 6695172.
- ↑ Thomson, M. M., Perez-Alvarez, L. and Najera, R. (2002). "Molecular epidemiology of HIV-1 genetic forms and its significance for vaccine development and therapy". Lancet Infect. Dis. 2 (8): 461–471. doi:. PMID 12150845.
- ↑ Carr, J. K.; Foley, B. T., Leitner, T., Salminen, M., Korber, B. and McCutchan, F. (1998). "Reference Sequences Representing the Principal Genetic Diversity of HIV-1 in the Pandemic". in Los Alamos National Laboratory (ed.) (PDF format). HIV Sequence Compendium. Los Alamos, New Mexico: Los Alamos National Laboratory. pp. 10–19.
- ↑ Osmanov S, Pattou C, Walker N, Schwardlander B, Esparza J; WHO-UNAIDS Network for HIV Isolation and Characterization. (2002). "Estimated global distribution and regional spread of HIV-1 genetic subtypes in the year 2000". Acquir. Immune. Defic. Syndr. 29 (2): 184–190. PMID 11832690.
- ↑ Perrin L, Kaiser L, Yerly S. (2003). "Travel and the spread of HIV-1 genetic variants". Lancet Infect Dis. 3 (1): 22–27. doi:. PMID 12505029.
- ↑ Piatak, M., Jr, Saag, M. S., Yang, L. C., Clark, S. J., Kappes, J. C., Luk, K. C., Hahn, B. H., Shaw, G. M. and Lifson, J.D. (1993). "High levels of HIV-1 in plasma during all stages of infection determined by competitive PCR". Science 259 (5102): 1749–1754. doi:. PMID 8096089.
- ↑ Pantaleo G, Demarest JF, Schacker T, Vaccarezza M, Cohen OJ, Daucher M, Graziosi C, Schnittman SS, Quinn TC, Shaw GM, Perrin L, Tambussi G, Lazzarin A, Sekaly RP, Soudeyns H, Corey L, Fauci AS. (1997). "The qualitative nature of the primary immune response to HIV infection is a prognosticator of disease progression independent of the initial level of plasma viremia". Proc Natl Acad Sci U S A. 94 (1): 254–258. doi:. PMID 8990195.
- ↑ Kahn, J. O. and Walker, B. D. (1998). "Acute Human Immunodeficiency Virus type 1 infection". N. Engl. J. Med. 331 (1): 33–39. doi:. PMID 9647878.
- ↑ Daar ES, Little S, Pitt J, et al (2001). "Diagnosis of primary HIV-1 infection. Los Angeles County Primary HIV Infection Recruitment Network". Ann. Intern. Med. 134 (1): 25–9. PMID 11187417.
- ↑ Burton GF, Keele BF, Estes JD, Thacker TC, Gartner S. (2002). "Follicular dendritic cell contributions to HIV pathogenesis". Semin Immunol. 14 (4): 275–284. doi:. PMID 12163303.
- ↑ Clapham PR, McKnight A. (2001). "HIV-1 receptors and cell tropism". Br Med Bull. 58 (4): 43–59. doi:. PMID 11714623.
- ↑ 95.0 95.1 95.2 95.3 Kumaranayake, L. and Watts, C. (2001). "Resource allocation and priority setting of HIV/AIDS interventions: addressing the generalized epidemic in sub-Saharan Africa". J. Int. Dev. 13 (4): 451–466. doi:.
- ↑ Kleinman, S. (September 2004). "Patient information: Blood donation and transfusion". Uptodate. Retrieved on 2008-04-03.
- ↑ 97.0 97.1 Centers for Disease Control and Prevention. (2001). "Revised guidelines for HIV counseling, testing, and referral". MMWR Recomm Rep. 50 (RR-19): 1–57. PMID 11718472.
- ↑ Celum CL, Coombs RW, Lafferty W, Inui TS, Louie PH, Gates CA, McCreedy BJ, Egan R, Grove T, Alexander S, et al. (1991). "Indeterminate human immunodeficiency virus type 1 western blots: seroconversion risk, specificity of supplemental tests, and an algorithm for evaluation". J Infect Dis. 164 (4): 656–664. PMID 1894929.
- ↑ Robb ML. "Failure of the Merck HIV vaccine: an uncertain step forward". Lancet 372 (9653): 1857–1858. doi:.
- ↑ Fan, H., Conner, R. F. and Villarreal, L. P. eds, ed. (2005). AIDS : science and society (4th edition ed.). Boston, MA: Jones and Bartlett Publishers. ISBN 0-7637-0086-X.
- ↑ Department of Health and Human Services (January 2005). "A Pocket Guide to Adult HIV/AIDS Treatment January 2005 edition". Retrieved on 2006-01-17.
- ↑ 102.0 102.1 Palella, F. J., Delaney, K. M., Moorman, A. C., Loveless, M. O., Fuhrer, J., Satten, G. A., Aschman, D. J. and Holmberg, S. D. (1998). "Declining morbidity and mortality among patients with advanced human immunodeficiency virus infection". N. Engl. J. Med. 338 (13): 853–860. doi:. PMID 9516219.
- ↑ Department of Health and Human Services Working Group on Antiretroviral Therapy and Medical Management of HIV-Infected Children (November 3, 2005). "Guidelines for the Use of Antiretroviral Agents in Pediatric HIV Infection" (PDF). Retrieved on 2006-01-17.
- ↑ Department of Health and Human Services Panel on Clinical Practices for Treatment of HIV Infection (October 6, 2005). "Guidelines for the Use of Antiretroviral Agents in HIV-1-Infected Adults and Adolescents" (PDF). Retrieved on 2006-01-17.
- ↑ Martinez-Picado, J., DePasquale, M. P., Kartsonis, N., Hanna, G. J., Wong, J., Finzi, D., Rosenberg, E., Gunthard, H.F., Sutton, L., Savara, A., Petropoulos, C. J., Hellmann, N., Walker, B. D., Richman, D. D., Siliciano, R. and D'Aquila, R. T. (2000). "Antiretroviral resistance during successful therapy of human immunodeficiency virus type 1 infection". Proc. Natl. Acad. Sci. U. S. A. 97 (20): 10948–10953. doi:. PMID 11005867.
- ↑ Dybul, M., Fauci, A. S., Bartlett, J. G., Kaplan, J. E., Pau, A. K.; Panel on Clinical Practices for Treatment of HIV. (2002). "Guidelines for using antiretroviral agents among HIV-infected adults and adolescents". Ann. Intern. Med. 137 (5 Pt 2): 381–433. PMID 12617573.
- ↑ Blankson, J. N., Persaud, D., Siliciano, R. F. (2002). "The challenge of viral reservoirs in HIV-1 infection". Annu. Rev. Med. 53: 557–593. doi:. PMID 11818490.
- ↑ 108.0 108.1 Wood, E., Hogg, R. S., Yip, B., Harrigan, P. R., O'Shaughnessy, M. V. and Montaner, J. S. (2003). "Is there a baseline CD4 cell count that precludes a survival response to modern antiretroviral therapy?". AIDS 17 (5): 711-720. PMID 12646794.
- ↑ Chene, G., Sterne, J. A., May, M., Costagliola, D., Ledergerber, B., Phillips, A. N., Dabis, F., Lundgren, J., D'Arminio Monforte, A., de Wolf, F., Hogg, R., Reiss, P., Justice, A., Leport, C., Staszewski, S., Gill, J., Fatkenheuer, G., Egger, M. E. and the Antiretroviral Therapy Cohort Collaboration. (2003). "Prognostic importance of initial response in HIV-1 infected patients starting potent antiretroviral therapy: analysis of prospective studies". Lancet 362 (9385): 679-686. PMID 12957089.
- ↑ A computer based study in 2006, following the 2004 United States treatment guidelines: Schackman BR, Gebo KA, Walensky RP, Losina E, Muccio T, Sax PE, Weinstein MC, Seage GR 3rd, Moore RD, Freedberg KA. (2006). "The lifetime cost of current HIV care in the United States". Med Care 44 (11): 990-997. PMID 17063130.
- ↑ Becker SL, Dezii CM, Burtcel B, Kawabata H, Hodder S. (2002). "Young HIV-infected adults are at greater risk for medication nonadherence". MedGenMed. 4 (3): 21. PMID 12466764.
- ↑ Nieuwkerk, P., Sprangers, M., Burger, D., Hoetelmans, R. M., Hugen, P. W., Danner, S. A., van Der Ende, M. E., Schneider, M. M., Schrey, G., Meenhorst, P. L., Sprenger, H. G., Kauffmann, R. H., Jambroes, M., Chesney, M. A., de Wolf, F., Lange, J. M. and the ATHENA Project. (2001). "Limited Patient Adherence to Highly Active Antiretroviral Therapy for HIV-1 Infection in an Observational Cohort Study". Arch. Intern. Med. 161 (16): 1962-1968. PMID 11525698.
- ↑ Kleeberger, C., Phair, J., Strathdee, S., Detels, R., Kingsley, L. and Jacobson, L. P. (2001). "Determinants of Heterogeneous Adherence to HIV-Antiretroviral Therapies in the Multicenter AIDS Cohort Study". J. Acquir. Immune Defic. Syndr. 26 (1): 82-92. PMID 11176272.
- ↑ Heath, K. V., Singer, J., O'Shaughnessy, M. V., Montaner, J. S. and Hogg, R. S. (2002). "Intentional Nonadherence Due to Adverse Symptoms Associated With Antiretroviral Therapy". J. Acquir. Immune Defic. Syndr. 31 (2): 211-217. PMID 12394800.
- ↑ Montessori, V., Press, N., Harris, M., Akagi, L., Montaner, J. S. (2004). "Adverse effects of antiretroviral therapy for HIV infection.". CMAJ 170 (2): 229-238. PMID 14734438.
- ↑ Saitoh, A., Hull, A. D., Franklin, P. and Spector, S. A. (2005). "Myelomeningocele in an infant with intrauterine exposure to efavirenz". J. Perinatol. 25 (8): 555-556. PMID 16047034.
- ↑ Wang C, Vlahov D, Galai N, et al. (2004). "Mortality in HIV-seropositive versus seronegative persons in the era of highly active antiretroviral therapy.". J. Infect. Dis. 190: 1046–54. PMID 15319852.
- ↑ World Health Organisation (2006). "WHO case definitions of HIV for surveillance and revised clinical staging and immunological classification" (PDF). Retrieved on 2006-12-27.
- ↑ 119.0 119.1 119.2 Ferrantelli F, Cafaro A, Ensoli B. (2004). "Nonstructural HIV proteins as targets for prophylactic or therapeutic vaccines". Curr Opin Biotechnol. 15 (6): 543-556. PMID 15560981.
- ↑ Researchers at the Heinrich Pette Institute of Experimental Virology and Immunology at Hamburg:Paternity Testing Labs (2007). "German scientists “cure” HIV-infected human lymphocytes". Retrieved on 2008-04-03.
- ↑ scientists at the Max Planck Institute for Molecular Cell Biology and Genetics in Dresden: Terra Daily (2007). "Another Potential Cure For HIV Discovered". Retrieved on 2007-06-29.
- ↑ 122.0 122.1 Mark Schoofs (2008). "A Doctor, a Mutation and a Potential Cure for AIDS". The Wall Street Journal. Retrieved on 2008-11-09.
- ↑ UNAIDS, WHO (December 2007). "2007 AIDS epidemic update" (PDF). Retrieved on 2008-03-12.
- ↑ McNeil, Jr., Donald (November 20, 2007). "U.N. Agency to Say It Overstated Extent of H.I.V. Cases by Millions", The New York Times. Retrieved on 2008-01-16.
- ↑ UNAIDS (2001). "Special Session of the General Assembly on HIV/AIDS Round table 3 Socio-economic impact of the epidemic and the strengthening of national capacities to combat HIV/AIDS" (PDF format). Retrieved on 2006-06-15.
- ↑ World Bank (2005). "Evaluating the World Bank's Assistance for Fighting the HIV/AIDS Epidemic". Retrieved on 2006-01-17.
- ↑ Centers for Disease Control and Prevention (2002). "Cases of HIV Infection and AIDS in the United States, 2002". HIV/AIDS Surveillance Report. http://www.cdc.gov/hiv/topics/surveillance/resources/reports/2002report/table1.htm.
- ↑ Duesberg, P. H. (1988). "HIV is not the cause of AIDS". Science 241 (4865): 514, 517. doi:. PMID 3399880.
- ↑ Papadopulos-Eleopulos, E., Turner, V. F., Papadimitriou, J., Page, B., Causer, D., Alfonso, H., Mhlongo, S., Miller, T., Maniotis, A. and Fiala, C. (2004). "A critique of the Montagnier evidence for the HIV/AIDS hypothesis". Med Hypotheses 63 (4): 597–601. PMID 15325002.
- ↑ For evidence that HIV is the cause of AIDS, see (for example):
- , (2000). "The Durban Declaration". Nature 406 (6791): 15–6. doi:. PMID 10894520. - full text here.
- Cohen, J. (1994). "The Controversy over HIV and AIDS" (PDF). Science 266 (5191): 1642–1649. doi:. PMID 7992043. http://www.sciencemag.org/feature/data/cohen/266-5191-1642a.pdf.
- Various. "Focus on the HIV-AIDS Connection: Resource links". National Institute of Allergy and Infectious Diseases. Retrieved on 2006-09-07.
- O'Brien SJ, Goedert JJ (1996). "HIV causes AIDS: Koch's postulates fulfilled". Curr. Opin. Immunol. 8 (5): 613–8. PMID 8902385.
- Galéa P, Chermann JC (1998). "HIV as the cause of AIDS and associated diseases". Genetica 104 (2): 133–42. PMID 10220906.
- ↑ Watson J (2006). "Scientists, activists sue South Africa's AIDS 'denialists'". Nat. Med. 12 (1): 6. doi:. PMID 16397537.
- ↑ Baleta A (2003). "S Africa's AIDS activists accuse government of murder". Lancet 361 (9363): 1105. doi:. PMID 12672319.
- ↑ Cohen J (2000). "South Africa's new enemy". Science 288 (5474): 2168–70. doi:. PMID 10896606.
External links
- HIV/AIDS at the Open Directory Project
- "AIDSinfo - HIV/AIDS Treatment Information". US Department of Health and Human Services. Retrieved on 2008-03-21.
- "UNAIDS: The Joint United Nations Programme on HIV/AIDS". UNAIDS. Retrieved on 2008-03-21.
- "AIDS.gov: Portal to all Federal HIV/AIDS information". AIDS.gov. Retrieved on 2008-05-20.
- "HIV News and Resources". The Body. Retrieved on 2008-07-18.
|
|||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||