Fitness (biology)
Fitness (often denoted  in population genetics models) is a central concept in evolutionary theory. It describes the capability of an individual of certain genotype to reproduce, and usually is equal to the proportion of the individual's genes in all the genes of the next generation. If differences in individual genotypes affect fitness, then the frequencies of the genotypes will change over generations; the genotypes with higher fitness become more common. This process is called natural selection.
in population genetics models) is a central concept in evolutionary theory. It describes the capability of an individual of certain genotype to reproduce, and usually is equal to the proportion of the individual's genes in all the genes of the next generation. If differences in individual genotypes affect fitness, then the frequencies of the genotypes will change over generations; the genotypes with higher fitness become more common. This process is called natural selection.
An individual's fitness is manifested through its phenotype. As phenotype is affected by both genes and environment, the fitnesses of different individuals with the same genotype are not necessarily equal, but depend on the environment in which the individuals live. However, since the fitness of the genotype is an averaged quantity, it will reflect the reproductive outcomes of all individuals with that genotype.
As fitness measures the quantity of the copies of the genes of an individual in the next generation, it doesn't really matter how the genes arrive in the next generation. That is, for an individual it is equally "beneficial" to reproduce itself, or to help relatives with similar genes to reproduce, as long as similar amount of copies of individual's genes get passed on to the next generation. Selection which promotes this kind of helper behaviour is called kin selection.
The concept is particularly difficult to understand and frequently misunderstood; J.B.S. Haldane when discussing it with John Maynard Smith is reported to have described it as "a bugger".
Contents |
Measures of fitness
There are two commonly used measures of fitness; absolute fitness and relative fitness.
Absolute fitness (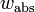 ) of a genotype is defined as the ratio between the number of individuals with that genotype after selection to those before selection. It is calculated for a single generation and may be calculated from absolute numbers or from frequencies. When the fitness is larger than 1.0, the genotype increases in frequency; a ratio smaller than 1.0 indicates a decrease in frequency.
) of a genotype is defined as the ratio between the number of individuals with that genotype after selection to those before selection. It is calculated for a single generation and may be calculated from absolute numbers or from frequencies. When the fitness is larger than 1.0, the genotype increases in frequency; a ratio smaller than 1.0 indicates a decrease in frequency.
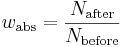
Absolute fitness for a genotype can also be calculated as the product of the proportion survival times the average fecundity.
Relative fitness is quantified as the average number of surviving progeny of a particular genotype compared with average number of surviving progeny of competing genotypes after a single generation, i.e. one genotype is normalized at 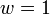 and the fitnesses of other genotypes are measured with respect to that genotype. Relative fitness can therefore take any nonnegative value, including 0.
and the fitnesses of other genotypes are measured with respect to that genotype. Relative fitness can therefore take any nonnegative value, including 0.
While researchers can usually measure relative fitness, absolute fitness is more difficult. It is often difficult to determine how many individuals of a genotype there were immediately after reproduction.
The two concepts are related, and both of them are equivalent when they are divided by the mean fitness, which is weighted by genotype frequencies.
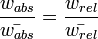
Because fitness is a coefficient, and a variable may be multiplied by it several times, biologists may work with "log fitness" (particularly so before the advent of computers). By taking the logarithm of fitness each term may be added rather than multiplied. A fitness landscape, first conceptualized by Sewall Wright, is a way of visualising fitness in terms of a three-dimensional surface on which peaks correspond to local fitness maxima; it is often said that natural selection always progresses uphill but can only do so locally. This can result in suboptimal local maxima becoming stable, because natural selection cannot return to the less-fit "valleys" of the landscape on the way to reach higher peaks.
The related concept of genetic load measures the overall fitness of a population of individuals of many genotypes whose fitnesses vary, relative to a hypothetical population in which the most fit genotype has become fixed.
As another example we may mention the definition of fitness given by Maynard Smith in the following way: ”Fitness is a property, not of an individual, but of a class of individuals – for example homozygous for allele A at a particular locus. Thus the phrase ’expected number of offspring’ means the average number, not the number produced by some one individual. If the first human infant with a gene for levitation were struck by lightning in its pram, this would not prove the new genotype to have low fitness, but only that the particular child was unlucky.” This measure is certainly useful in breeding programs, but hardly as a basis of a model of an evolution selecting individuals, because evolution would hardly know if the individual may be selected or not.
Yet another possible measure has been formulated by Hartl,1981: "The fitness of the individual - having an array x of phenotypes - is the probability, s(x), that the individual will be included among the group selected as parents of the next generation." Then, the mean fitness may be determined as a mean over the set of individuals in a large population.
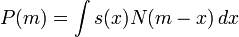
where N is the p. d. f. of phenotypes in the population, and m is its centre of gravity. This measure is a suitable basis of a model of an evolution selecting individuals. It may in principle take even the stroke of the lightning into consideration. In the case N is a Gaussian it is fairly easily proved that the average information (information entropy, disorder, diversity) of a large population may be maximized by Gaussian adaptation - keeping the mean fitness constant - in accordance with recapitulation, the central limit theorem, the Hardy-Weinberg law and the second law of thermodynamics. This is in contrast to Fisher's fundamental theorem of natural selection.
History
The British sociologist Herbert Spencer coined the phrase "survival of the fittest" (though originally, and perhaps more accurately, "survival of the best fitted") in his 1851 work Social Statics and later used it to characterise what Charles Darwin had called natural selection. The British biologist J.B.S. Haldane was the first to quantify fitness, in terms of the modern evolutionary synthesis of Darwinism and Mendelian genetics starting with his 1924 paper A Mathematical Theory of Natural and Artificial Selection. The next further advance was the introduction of the concept of inclusive fitness by the British biologist W.D. Hamilton in 1964 in his paper on The Evolution of Social Behavior.
Further reading
- Sober, E. (2001). The Two Faces of Fitness. In R. Singh, D. Paul, C. Krimbas, and J. Beatty (Eds.), Thinking about Evolution: Historical, Philosophical, and Political Perspectives. Cambridge University Press, pp.309-321. Full text
See also
- Gene-centered view of evolution
- Inclusive fitness
- Natural selection
- Reproductive success
- Selection coefficient